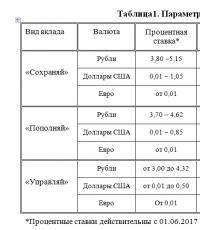
فاجعه کیسنف فاجعه کیسناوا در تاریخ زمین فاجعه کیسناوا در تاریخ زمین
در اقیانوس های اولیه و در جو زمین جوان ، هیچ ترشی وجود نداشت ، اگر آنها می خواستند فتوسنتز سیانوباکتری ها را تولید کنند و یک محصول جانبی متابولیسم تولید می کردند. ویلنی کیسن با عناصر در حال گسترش روی کره زمین مانند نیتروژن یا زغال سنگ در تعامل نیست ، اما محور افراد حیاتی است. در چند سال گذشته ، "ازدحام" زیادی از ترش ترش که در حدود سه میلیون سال پیش به زمین آمد و نزدیک به 2.4 میلیون سال پیش وجود داشت ، در آن زمان ترش در جو به سرعت افزایش یافت: در 200 میلیون ترشی بیش از 10 میلیون سال پیش افزایش یافته است! Tsya podіya prelіdniki تعمید داد فاجعه sisnevoy(رویداد اکسیداسیون بزرگ ، روشن. اکسیداسیون بزرگ) و ماهیت واکنشهای شیمیایی سطح زمین تغییر کرد.
انقلاب کیسناوا: چگونه زمین دوباره ایجاد شد
زمین شناس دانشگاه بریتیش کلمبیا متتیس اسمیت (Matthijs Smit) و همکارش ، پروفسور کلاوس مزگر (کلاوس مزگر) از دانشگاه برن ، یک ربات جدید را به تکامل این پدیده اختصاص دادند. شما می دانید که فاجعه کیسنف نژادهایی را که قاره ها از آن تشکیل شده اند نیز دگرگون کرد ، و اکنون آنها نتیجه تجزیه و تحلیل ژئوشیمیایی فعالیت آتشفشانی را در تمام نور تصور کردند ، اما در کیسه بی پایان ، 48000 نفر از آنها 48000 را مجاز کردند
Smit در بیانیه مطبوعاتی خود به این معنی است که از آن لحظه ، مانند اقیانوس ها ، مقدار زیادی بوسه وجود دارد ، در انبارهای قاره ها ، مارهای زیادی وجود دارد. نژادهای گیرسکی در قلمرو ایسلند فعلی و جزایر فارو در پشت انبار تقریباً مشابه نژادهای موجود در زمین جوان قبل از فاجعه کیسنووی است: بوی تعفن بسیاری از مغناطیس ها و محور سیلیس ستمکار در آنها کم است. نژاد گذشته معدنی است اولیوین، Yakiy іnіtsіyuvav kisnevі واکنش های شیمیایی در تماس با آب. در جهان ، همانطور که پوسته قاره ای در حال توسعه و بزرگ شدن بود ، اولیوین عملاً یک فرد آگاه است و با آن واکنش ها نیز انجام شد. کیسن در اقیانوس ها انباشته شده است و اگر آب فراوان بوده است ، گاز به جو جاری شده است.
به نظر می رسد که این یک نقطه مرجع برای توسعه اشکال زنده ای است که آنها شناخته می شوند. ترشي ديوانه كننده زمين نه تنها به طور كل به جايگاه عمومي زندگي تبديل شده است ، بلكه زيبا تر از آن به دنبال توسعه موجودات تاشو بوده است. دلیل تغییر ساختار قاره ای ناشناخته باقی مانده است ، اما این بدان معنا نیست که تکتونیک صفحه تقریباً در کل دوره بهبود یافته است و می توان بین آنها ارتباط مستقیم برقرار کرد.
مقدار نمایش
من در مورد تکامل و غیرزایی صحبت نمی کنم - تغذیه جوانه زنی روی زمین روی زمین هنوز نمایان تر است. با این حال ، بوسه عنصر بسیار مهمی است که زندگی کودک را حفظ می کند. می دانم که با تغییر زمین ، می توانم به همان اصل در مراحل مقدماتی دست یابم. سیاره های فراخورشیدیі در ارتعاش شدید برای مردم ، سیاره ای ایده آل برای استقرار: به عنوان مثال ، ستاره شناسان فوراً می بینند که دو سیاره در سیستم TRAPPIST-1 اقیانوس های باشکوه را پوشش می دهند. آگاهانه ، مانند سوشی که به شکل قاره ها تزریق شده است ، می توان صدای قابل توجهی از سر و صدا و صدای مستقیم حداکثر نور جدید را که به ما داده می شود ، شنید.
من شخصیت زاگالنی جو را از اصلی به حالت اکسیداتیو تغییر می دهم. اعتراف در مورد فاجعه Kisnev بر اساس رشد رشد شراب به ماهیت تجمع کالاهای افتاده شکسته شد.
اولین انبار اتمسفر
انبار دقیق جو اولیه زمین در روز ناراحتی امسال ، با این حال ، برای جایگزینی حال حاضر ، در نتیجه گوشته گاز زدایی شکل گرفت و ماهیت جدیدی داشت. اساس її توسط گاز دی اکسید کربن ، آب سر ، آمونیاک ، متان ساخته شد. در مورد دارچین سگو کاهش دهید:
- عدم اکسیداسیون ، که به وضوح روی سطح ایجاد شده است (به عنوان مثال ، یک سنگریزه کوچک از پودر ناپایدار تا ترش) ؛
- مشاهده گونه های قابل توجه ترشل ژرل و سایر اکسید کننده ها.
- vivchennya dzherels بالقوه جو اولیه (گازهای آتشفشانی ، انبار اولین اجرام آسمانی).
علل فاجعه ترش
یکی از عوامل مهم ترش مولکولی ، زیست کره و به طور دقیق تر موجودات فتوسنتزی است. فتوسنتز ، مبهم ، با ظهور در طلوع بیوسفر (3.7-3.8 میلیارد سال پیش) ، باستان شناسی و تعداد زیادی از گروههای باکتریایی در طول فتوسنتز موسلین نقض نشد. فتوسنتز Kisnevy از انگور در سیانوباکتریها 2.7-2.8 میلیارد روبل. Virіznyaytsya بوسه عملا در یک زمان vitrahavsya بر روی سنگهای girskikh اکسیده ، razchinennyh spoluk و گازهای جو. غلظت ویسوکا به صورت محلی ، در محدوده تشک های باکتریایی (t. sound. "Kisnevi kisheni") پخش شد. بدین منظور ، هم سنگهای سطحی و هم گازهای اتمسفر تبدیل به اکسیداسیون شده اند و هم رطوبت در بیابان در اتمسفر جمع شده است.
Bula یکی از قوی ترین عواملی است که در تغییر ناحیه میکروبی تحت تأثیر قرار گرفته است. انبار شیمیاقیانوس ، ویکلیکان فعالیت آتشفشانی
آثار فاجعه کیسنوو
زیست کره
Oskіlki بخش مهمی از موجودات زنده در آن ساعت ناآرامی زورگو ، ناشناخته غیر قابل توجه با غلظت قابل توجهی از ترش است ، تغییر جهانی روحیات در حال افزایش است: ارواح بیهوشی با هوازی تغییر کرده و با گونه های قبلی تجمع می کنند. ارواح بی هوازی ، ناوپاکی ، در "کیشنی بی هوازی" vitisneni ظاهر شدند (در ظاهر ، به نظر می رسد "زیست کره ناویوریت ظاهر شد"). ظاهر ترش مولکولی در اتمسفر منجر به تشکیل صفحه ای ازن شده و به طور قابل توجهی بند ناحیه بیوسفر را گسترش داده و دوقطبی ترش (از نظر انرژی) زنده تر (در برخی موارد بی هوازی) را به وجود آورده است.
جو
در نتیجه تغییر در شیمی جو فاجعه ترش ، تغییر در فعالیت شیمی ، ایجاد توپ ازن ، اثر گلخانه ای به طور چشمگیری تغییر کرده است. یاک به ارث برده ، این سیاره وارد عصر یخبندان هورون شد.
در مورد مقاله "فاجعه کیسناوا" پیام ارسال کنید
یادداشت
posilannya
- - طبیعت 458 ، 750-753 (09.04.2009)(انگلیسی)
- - CNews ، 03.08.2010
- نایمارک ، اولنا... elementy.ru (2.03.14). ...
|
||||||||||||||||||||||||||||||||||||||||||
Urivok ، که فاجعه Kisnev را مشخص می کند
نبرد بورودینسکی با ویسنوکی ، شکستن آن در اشغال مسکو و فرانسه ، بدون نبردهای جدید ، یکی از مهمترین رویدادهای تاریخ است.تمام تاریخ جهان در این واقعیت است که ندای قدرت قدرتها و ملتها ، در پیوندهای بین خودشان ، تغییر می کند. اما بدون تردید ، پس از موفقیت های بزرگتر و کمتر آن دوران ، قدرت سیاسی قدرتها و مردم تغییر خواهد کرد.
توصیفات تاریخی شگفت انگیزی در مورد آن نیست ، مانند یک پادشاه ، یا یک امپراتور ، که با آنامار ، یا یک پادشاه جوش داده و مردم را در کیلک میلیون یورو تحت تسلط خود درآورده است. از آنجا که منطقی نبود ، چرا شکست یک ارتش ، یکی از همه نیروهای مردم ، مردم عصبانی بودند ، همه پذیرفته شده بودند ، صدها نشانه بهبود یا تغییر در قدرت مردم. این تغییر بسیار عالی بود و حق تغییر افراد به Skoda جدید بهبود یافت. وایسکو از شلیک ها مطلع بود ، و بی گناه است ، پس از مرحله نبرد ، به مردم اجازه داده می شود که راست بگویند ، و وقتی به سرعت به ضربات خود ضربه می زنند ، آماده شروع خواهند بود.
بنابراین (از تاریخ) از ساعات پیدا شده و تا ساعت کنونی بود. همه هشدارهای ناپلئون باید ضامن این قاعده باشند. در پشت مرحله مبارزه پیروزی های اتریش - اتریش آزادی حقوق خود و بهبود حقوق و قدرت فرانسه است. غلبه بر فرانسوی ها توسط ایوان و اورشتت به معنای استقلال پروس است.
در سال 1812 ، در سال 1812 ، فرانسوی ها به مسکو پیروز شدند ، مسکو به تصرف درآمد ، و در کنار تیم ، بدون نبردهای جدید ، روسیه تسلط خود را متوقف نکرد ، اما به جای فرانسه ناپلئون ، از به دست آوردن یک ارتش ششصد هزار نفری دست کشید. حقایق مربوط به قواعد تاریخ را گسترش دهید ، بگویید که میدان نبرد در بورودینو تحت الشعاع روسها قرار گرفت ، و مسکووی آنها را به جنگ فرستاد ، زیرا ارتش ناپلئون را خراب کردند ، حیف نیست.
پیروزی های پیسلیا بورودین در فرانسوی ها نه تنها به طور کلی ، بلکه به طور کلی نبرد مهمی را به همراه نداشت و ارتش فرانسه دیگر کشیده نشد. چه مفهومی داره؟ من می توانم بگویم که این پدیده تاریخی نیست (اگر از جهان نشأت نگیرد ، تنبل تاریخ است). همانطور که در سمت راست ، ما می توانیم ترفند غیر پیش پا افتاده را انتخاب کنیم ، که در آن گلوله کمی عقب مانده است ، می توانیم آن را برای روشن شدن انتخاب کنیم. ale the podіya tse در چشم پدران ما دیده شد ، برای کسانی که تغذیه زندگی و مرگ زندگی ، و زندگی کل زندگی را از همه رزمندگان می دیدند ...
دوره کارزار 1812 از نبرد بورودین تا شکست فرانسوی ها بود ، اما این نبرد تنها به این دلیل انجام نشد که دلیل فتح نبود ، اما نشانه فتح نبود. doviv ، چه نیرویی ، به عنوان سهم مردم ، در فاتح نیست ، تا در ارتش و نبردها ادامه دهد ، بلکه در چیز دیگری است.
مورخان فرانسوی ، اردوی بازدیدکنندگان فرانسوی را قبل از خروج از مسکو توصیف می کنند ، همه چیز را برای ارتش بزرگ ، از جمله سواره نظام ، توپخانه و حمل و نقل ، که علوفه ای برای تغذیه اسب ها و اسب های نازک شاخدار ندارند ، در نظم نگه می دارند. نمی توان به هیچ وجه به این جرات کمک کرد ، بنابراین مردان ناوکولیشی گناهان خود را برداشته و به فرانسوی ها ندادند.
نبرد نتایج فوق العاده ای به همراه نداشت ، بنابراین مردان کارپو و ولاس ، که با کمک فرانسوی ها به مسکو آمده بودند ، با رانندگان گرابواتی به مسکو آمدند ، اما آنها به ویژه قهرمانانه ظاهر نشدند ، و من چنین نکردم. چنین مردان خوبی ندارید.
بدیهی است که دو نفر با رعایت تمام قوانین شاهکار شمشیربازی با شمشیر به دوئل رفته اند: حصارکشی برای رسیدن به آخرین ساعت بی اهمیت بود. با دیدن اینکه ما صدمه دیده ایم ، یکی از مخالفان را لرزاند-ما صدا می کردیم ، اما در سمت راست ، گرم نبود ، اما از این زندگی خجالت زده بود ، شمشیر خود را انداخت і ، لوماک اول و دوم را گرفت ، سعی کرد بچرخد تمام شد افسوس ، بدیهی است که دشمن به طرز زیرکانه ای زنده و درخشان است و به سادگی برای رسیدن به نقطه ، در عین حال با کمک دستورات مردم ، می خواهد به اصل موضوع تعصب داشته باشد و بی را تحت فشار قرار دهد. همه قوانین تغییر رمز و راز ممکن است مانند یک سرکش آگاه شوید ، و ابهام ناشی از چنین توصیفی از واقعیت است که به صورت اتفاقی تبدیل شده است.
شمشیر زن ، مانند ویماژ در مبارزه برای قوانین هنر ، بولز فرانسوی است. حریف شما ، پرتاب شمشیر و گرفتن چماق ، قلدر روسی ؛ افرادی که می خواهند همه چیز را در مورد قوانین شمشیربازی توضیح دهند ، مورخانی هستند که در مورد طب سوزنی نوشته اند.
در ساعت یک ، اسمولنسکا آماده رفتن بود ، اما نه برای جنگ. اتاق خواب شهر و سیل ، ورودی نبرد ، ضربه بورودین و ورودی جدید ، مازاد و آتش مسکو ، گرفتن غارتگران ، تغییر وسایل نقلیه ، جنگ پارتیزانها - همه اهداف قوانین.
به طور کلی پذیرفته شده است که اتمسفر مولکولی جو زیست زا است و کمربند دوم لزوماً با فرمولاسیون نوع جدیدی از فتوسنتز مرتبط نیست ، در حالی که آب در نقش اهدا کننده الکترون ، جانشین است. در ذهن زمین اولیه ، قبل از اعلام ، آنها صدف یوباکتریهای فتوسنتز کننده را با یک واکنش dzherel vіlnogo sisny bula به فوتولیز بخار آب در جو مشاهده می کنند ، همانطور که در زیر ماوراء بنفش دم کوتاه رخ می دهد. با این حال ، تعداد ترش "فوتولیتیک" ناچیز بود. تصویب بوسیله گازهای اکسید شده در جو اولیه و مواد معدنی تجدید شده ، باید وارد انبار سرخک زمینی شود.
ابتدایی ترین موجودات و همچنین فتوسنتز با تصاویر O2 ، اولیه ترین موجودات ، باکتری های فتوسنتز کننده (سیانوباکتری ها ، پروکلروفیت ها) و موجودات مناسب ، اما ظاهر اکسیژنات مولکولی
تا کشف یوکارهای فتوسنتزی و در پرشو چرگو ویشیخ رزلینتغییر ترش زنده در جو زمین در زمینه جو مدرن ناچیز است. با این حال ، پس از انجام پدراخانک ، برای مخلوط کردن ارگانیسم از سرگردان به سطح انرژی ، غلظت 0.2 enough کافی است ، یعنی سطح 0.01 در یک فضای مدرن. ظهور شمع O2 در جو زمین رخ می دهد ، به این معنی که ارزیابی مجدد برای زندگی بعدی روی زمین مهم است. perch for all tse به معنی صد قریب به اتفاق همه چیز بود ، که در دوران "قبل از اکسیژن" روی زمین فرموله شد و در وهله اول ، موجودات زنده در حال رشد بودند.
Osvita O2 در مقادیر زیاد جریان واکنشهای اکسیداتیو را در مقیاس بزرگ کاهش می دهد. تغییر ماهیت جو: از vona vodnovlyuvalnoy اکسید شد. بقیه تغییرات زیادی را در مشکل اهدا کننده و پذیرنده ایجاد کرده است. در حالی که در ذهن یک جو عاری از اسید تحت تسلط راه حل مشکل گیرنده الکترون قرار دارد ، در ذهن یک فضای ترش مشکل اصلی اهدا کننده الکترون هنوز مشکل اصلی است ، زیرا ظاهر O2 است در جو زمین ، معجزه پذیرنده الکترونها ایجاد شد.
اتصال PROCARIOTIV 3 بوسه مولکولی
مشتی صدف مولکولی در وسط سلول ها ظاهر شد و بلافاصله مشکل تعامل بین سلول های O2 در اتاق را حل کرد. آنها همچنین از اشکال مختلف زندگی خلاص نشدند. این می تواند یک کمک هزینه باشد ، که اولین نوع تعامل با O2 است که بر اساس نگرش بسیار منفی به کلیتینی جدید است. کاربرد داده های کلی - عددی در مورد سمیت بالای اسید مولکولی برای ضروری ترین موجودات بی هوازی 62.
62 با حیله و تزویر دانک سیکاوی در مورد کسانی که در دوره ای که مقدار زیادی ترش در جو وجود دارد ، درست در آغاز ساعت ورزش ورزشکاران طرفدار کاروتاسیون همه کاره تر می شود. تنوع واگن های طرفدار به میزان قابل توجهی 1.5 میلیارد روبل تغییر کرده است (شکل 52).
در جهان ، تجمع O2 به یک جزء پایدار آخرین طبقه متوسط تبدیل می شود و فقط در سطح محلی می توان چنین پیامدهایی را ایجاد کرد ، که به صورت روزمره اتفاق می افتد یا در مقدار کمی اتفاق می افتد. دو گزینه ممکن برای تعامل پروکاروئیدها با اسید مولکولی وجود دارد. برخی از اشکال مختلف بی هوازی "به محل زندگی" رفتند ، de O 2 عملاً قابل مشاهده است ، و ما به خودی خود "چشم انداز دوران بدون اسید" را نجات دادیم. інші دسته از zmushenі نوشیدن در طول راه pristosuvannya به ذهن "ترش". این بدان معناست که بوی بد واکنشهای متابولیکی جدیدی را شکل می دهد و در وهله اول برای خنثی سازی ترشح مولکولی منفی تزریق شده عمل می کند.
Otzhe ، قلاب دوزی آینده در راه ارتباط بین پروکاریا و ترش - قدرت حضور در حضور O 2 ، خنثی کردن این عمل منفی. با آواز خواندن خارج از سیستم ، شکل گیری ترش مولکولی در پروکاروت ها را می توان حذف کرد ، نمایندگان گروه را می توان حذف کرد و آنها را می توان بر روی درختان جوان درابینی تکاملی برشته کرد.
اثرات سمی اسیدیته مولکولی و سن
به عنوان عامل آخرین مرحله میانی ، O2 به دو طریق روی موجودات زاینده فعلی تزریق می شود: از یک طرف ، ممکن است کاملاً ضروری باشد ، از طرف دیگر - از ترش مولکولی و اثرات سمی مرتبط با قدیمی برای کلیتین ها.
بوسه مولکولی.تعدادی از فرضیه ها برای روشن شدن حساسیت پروکاروت ها تا O2 وجود دارد. بدیهی است که یکی از آنها ، اسید مولکولی به خودی خود سمی است ، که به دلیل توسعه عملکردهای اکسید کننده سلول ها برای نیاز ، تهاجمی است. اثر سمی O2 در همه ذهنها ذخیره می شود ، که موجودات زنده با آنها در تعامل هستند: غلظت O2 پراکنده ، بی اهمیتی نمایشگاه ، انبار وسط navkolishny.
به عنوان مثال ، سمیت منبع مولکولی می تواند استقبال زیادی از دستگاه های الکترونیکی از حامل های تلفن همراه باشد ، به طوری که در صورت سرگردانی ، در صورتی که منجر به تجمع نوع سلول داخلی برای تجدید اهداکنندگان شود ، عمل می کند. Dysno ، مشخص شده است که فعالیت فلاوپروتئین های رزین ، عملکردهای NAD (F) -H2 -اکسیداز را در طول رشد 5 -6 بار تغییر داده است. Clostridium acetobutylicumدر ذهن هوازی ، به نوعی از بی هوازی. تخریب حامل های الکترونیکی تزریق شده O 2 در جهت انتقال به آسیاب اکسیده شده ، که منجر به رشد و تغییر روش تولید در تخمیر می شود: افزایش سنتز اسید بوتیریک و تجمع اسید اکسید شده بیشتر -
نارشتی ، برای تجلی اثر سمی O2 ، اکسیداسیون کافی یک متابولیت کلیدی یا آنزیم وجود دارد که باید به نقطه فعال شدن برسد. سه سیستم آنزیمی و پروکاروئیدها ، به ویژه به اسیدیته مولکولی حساس هستند: نیتروژناز ، هیدروژناز و ریبولوز فسفات کربوکسیلاز.
سیستم نیتروژناز ، که تثبیت نیتروژن مولکولی را کاتالیز می کند ، در دو پروتئین فلزی ذخیره می شود: یک بطری ، که می تواند جایگزین طلا و مولیبدن شود و یک بطری ، در انباری که فقط طلا می تواند در آن قرار گیرد. به بلوک چرمی برای فعالیت کاتالیزوری سالم ضروری است. بوسه مولکولی به جرم نیتروژناز بسیار حساس است ، نسبت به بلوکهای O2 Fe کمی حساس تر است.
حساسیت نیتروژنازها تا O2 دلیل اصلی همه حساسیت مراکز فلزی است که در بستر حساس و همچنین در وسایل الکترونیکی منتقل شده شرکت می کنند. نوسانات در طیف وسیعی از مراحل ارتقاء O2 با مکانیسم تک الکترونیکی ، مانند محصولات چنین ارتقاء ، سوپراکسید ، پراکسید آب و صدف تک است که می تواند به اکسیداسیون اضافه شود.
پروتئین های نیتروژناز یک جزء واحد از سیستم تثبیت کننده نیتروژن نیستند ، نسبت به O2 حساس هستند. فردوکسین و فلاودوکسین ، که برق را به نیتروژناز اهدا می کنند ، می توانند خود اکسید شده و تبدیل به اکسیداتیو غیر عروقی شوند.
هیدروژنازهای پروکاریوت های باگاتوک نیز حساسیت بالایی به ترش مولکولی نشان می دهند ، همانطور که در شرایط آزمایشگاهی در جهان بزرگ طبق روش بینایی و مرحله تصفیه تعیین می شود. به عنوان یک قاعده ، مراحل بیشتری برای آماده سازی آنزیم خام O2 وجود دارد. در مورد آنزیم متصل به غشا ، قدرت تا O2 هیدروژناز ، از غشای چسبنده ، کمتر است. آنزیم ، otrimaniy از بی هوازی کلیتین ، حساس تر به O2 ، دید کمتر از پروکاروت های هوازی کلیتین.
فعالیت کاتالیزوری ریبولوز فسفات کربوکسیلاز ، آنزیمی که تثبیت کننده CO 2 در اکثریت قریب به اتفاق پروکاروتهای اتوتروف است ، برای رسوب بخشی از CO2 و O 2 . با غلظت بالای O2 و کم - CO2 ، واکنش اکسیژناز تبدیل می شود:
مولکولهای O2 و CO2 برای مرکز کاتالیزوری آنزیم رقابت می کنند. من می خواهم آنزیم خود به ترش مولکولی حساسیت ایجاد نکند و همچنین ، در غلظت بالا ، O2 را در وسط افزایش دهد تا عملکرد ریبولوزودی فسفات کربوکسیلاز کاهش یابد. واکنش آنزیمی بیش از حد در طول مسیر اکسیژناز برای تولید مجموعه ای از مولکولهای ریبولوز فسفات تا لخته شدن سلول ، به عنوان میراث کاهش فعالیت چرخه اصلی پنتوز فسفات در سلول.
تشخیص واکنشهای بیولوژیکی و عوامل فیزیکی و شیمیایی مختلف در شکل اولیه بسیار مهم است که محصولات نوآوری O2 نامناسب را شناسایی کنند ، که برای طبقه سالمتر از نظر واکنش پذیری سمی تر است. Yak vidomo ، برای معرفی مجدد اسیدیته مولکولی ، لازم است یک مولکول آب تولید شود تا زمانی که تأیید شود ، 4 الکترون مورد نیاز است:
O 2 + 4H + + 4 ه- H 2H 2 O
در تعداد زیادی از آنزیم های پروکاریوت ، که واکنش انتقال یک ساعته 4 الکترون به O2 را کاتالیز می کند ، که هیچ واسطه ای از O2 برای آنها تشخیص داده نمی شود. با این حال ، ممکن است محصولات کوتاه مدت تجدید ناقص O2 در این واکنش ها یافت شوند ، یا ممکن است با آنزیم ها گیر کرده ، وارد سیتوپلاسم نشوند و عملاً مکتبی از مشتریان راه اندازی نکنند.
آنیون سوپراکسیداگر تصویر مولکولی مرحله به مرحله به روز شود ، پس از انتقال 1 الکترون به O2 ، یک آنیون سوپراکسید (سوپراکسید) ایجاد می شود:
O 2+ ه- O 2 -.
انتقام گرفتن از الکترونهای جفت نشده را که با یک رادیکال (آنیون رادیکال) منفی است ، متوقف کنید. Win را می توان با فرمول یک رادیکال خنثی هیدروپراکسید خاموش کرد:
O 2 -. + H + HO 2.
در باقی ساعت ، viznannya این دیدگاه را رد کرد ، اما در اینجا امنیت اولیه ارگانیسم ها برای ارائه محصولاتی است که می توانند با یک مولکول تک الکترونی O2 ایجاد شوند. , یکی از آنها آنیون سوپراکسید است.
شما می توانید بسیاری از واکنش های بیوشیمیایی را که می توانند قبل از سالگرد ایجاد شوند ، نام ببرید. سوپر اکسید آنیونی در اثر برهم کنش با مولکولهای O2 سایر اجزا (تجدید فلاوین ، شینونی ، مرکاپتان ، پروتئین FeS) و همچنین در واکنشهایی که آنزیمهای فلاوپروتئین را کاتالیز می کنند ، تولید می شود. نارشتی ، در فرآیند فتوسنتز ، استفاده چندانی از الکترون ها وجود ندارد. واکنش های بیشتر به فتوسنتز - زنجیره واکنش به انتقال تک الکترون. آنیون سوپراکسید اغلب در سیستم یافت می شود. واکنش های کریم با طبیعت بیولوژیکی O2 -. هنگامی که با سونوگرافی تزریق می شوید ، در نتیجه فرایندهای فتوشیمیایی ، شیمیایی و الکتریکی ، می توانید حالت جیرجیرک را در سطح آب احساس کنید.
فعال بودن به صورت واکنشی به معنای جهان برای ماندن در ثبات ایمن نیست. به طور کلی ، آنها O2 هستند -. Velma ایمن نیست ، زیرا ساعت "زندگی" در محیط آبکی بی اهمیت است ، نه برای رادیکالهای مشتق از O2. به این ترتیب ، O2 - برونزا تعیین شد. ممکن است به داخل سلول (به ترتیب درون زا) نفوذ کرده و در واکنشها شرکت کنید ، که می تواند منجر به ایجاد اسیدهای چرب شود: پراکسیداسیون اسیدهای چرب اشباع نشده ، گروه های SH پروتئین های اکسیده شده ، DNA و مسافرخانه. سمیت آنیونهای سوپراکسید را می توان برای واکنشهای ثانویه افزایش داد که منجر به تأیید رادیکالهای هیدروکسی (OH) و اسید منفرد (* O 2) می شود.
بسیاری از پروکاریوتها ، که می توانند به سایر گروههای فیزیولوژیکی ارجاع داده شوند ، از جمله بی هوازی شدید ، ممکن است یک سرکوب کننده خاص برای آنزیم سوپراکسید دیسموتاز داشته باشند ، که یک بار اضافی سالم از یونهای O2 -است. و کاتالیزاسیون تحریف ها. آنیون سوپراکسید تأیید شده در واکنشی که خود به خود (3) یا کاتالیزور شده توسط سوپراکسید دیسموتاز (4) متلاشی می شود:
O 2 -. + O 2 -. + 2H + H 2 O 2 + * O 2 ؛ (3)
O 2 -. + O 2 -. + 2H + H 2 O 2 + O 2. (4)
ملاحظات واکنشهای این محصولات (فراوانی واکنش آنزیمی تقریباً به ترتیب بزرگی است ، نه خود به خود) ، و همچنین ، در صورت واکنش خود به خودی تغییر شکل ، یکی از تخمیرهای واکنش اغلب در ایستگاه سه گانه اصلی
رادیکال هیدروکسی Superoxidanion می تواند با H2O2 با فرمولاسیون رادیکال هیدروکسی (OH.) که O2 را نادیده می گیرد ، ارتباط داشته باشد. با فعالیت اکسیداتیو و سمیت:
O 2 -. + H 2 O 2 + H + O 2 + H 2 O + OH. ... (5)
Dzherel viniknennya OH. این می تواند واکنش اکسیداسیون تک الکترون با پراکسید آب را ارائه دهد ، مسیرهای گالوانیزه را که در سلولها مشهود است کاتالیز کند:
H 2 O 2 + Fe 2+ Fe 3+ + OH - + OH. ... (6)
علاوه بر واکنشهای قابل توجه رادیکالهای هیدروکسید ، آنها همچنین در مورد آب رادیواکتیو و در غلظتهای کم ، در سطح آب رخ می دهند. اوه از بهترین انواع اکسیداسیون ، قوی ترین ، اما استفاده از رادیواکتیویته انواع باگاتو از پلیمرهای زیستی.
پراکسیدانتقال 2 الکترون به O2 برای تولید آنیون پراکسید (7) یا پراکسید آب (8):
O 2 + 2 ه- O 2 2- ؛ (7)
O 2 + 2H + + 2 ه- H 2 O 2. (8)
کاتالیزور انتقال 2 الکترون به O2 را می توان در سلولهای پروکاریوتهای طبیعت اکسیداز فلاوین و دیاکی سیتوکروم یافت. Dzherelom H 2 O 2 می تواند واکنش خود اکسیداسیون برخی از پروتئینهای FeS غیر هم ، و همچنین توصیف واکنش تغییر شکل رادیکالهای سوپراکسید (واکنشهای 3 و 4) باشد. پراکسید در تمام آئروسل ها و بی هوازی های اختیاری وجود دارد ، بنابراین در ذهن هوایی رشد می کند ، بنابراین این یک فرایند طبیعی است.
آب پراکسید پایدارترین محصول صنعتی تجدید O2 ، ale و جدیدترین واکنش است. در تعداد زیادی پروکروت هوازی H2O2 ، آنها به سرعت در پشت افزودن آنزیم های حاوی هم ، کاتالاز و پراکسیداز قرار می گیرند. بدون آنها ، H2O2 می تواند در غلظت هایی که برای بدن کشنده هستند جمع شود.
H2O2 viclikє اکسیداسیون گروههای SH در سفیدها ، اکسیداسیون پراکسید اسیدهای چرب اشباع نشده. با این حال ، تعداد واکنشها برخلاف مواد vimirny ، مانند غلظت H 2 O 2 در سلولها ، برای چند مرتبه در اندازه ای که می خواهید به صورت in vivo به آن برسید. این چنین نیست ، زیرا پراکسید نه از طریق تعامل مستقیم با اجزای سلول ، بلکه با O2 واکنش پذیر است. (واکنش 5) با یونهای Fe2+ (واکنش 6) ، ممکن است برای تأیید رادیکال هیدروکسی تولید شود.
در دهه 20 این نظریه که سمیت O2 تجمع یافته در آب آب در سلول ها را توضیح می دهد ، بسیار محبوب بود. با این حال ، O2 سمی تر برای سلول در وسط محصولات اولیه و ثانویه اولین به روز رسانی تک الکترونیکی (O2 ، OH. * O2) یافت شد.
تک بوسه.هنجار O2 در یک اردوگاه پایدار است ، سه ساله نامیده می شود و با کمترین سطح انرژی مولکولی مشخص می شود. در ذهن خواننده ، مولکول O2 در یکی از دو stan_s (* O 2) شکسته منتقل می شود ، بنابراین با سطح انرژی و بی اهمیتی "زندگی" توسعه می یابد. در تعداد زیادی از سلولهای زنده در temryavi ، dzherel اصلی اسید منفرد ، تبدیل خود به خود آنیونهای سوپراکسید است (واکنش 3). بوسیله منفرد را نیز می توان با تعامل دو رادیکال تشخیص داد:
O 2 -. + اوه OH - + * O 2. (9)
اما ، این یک سیستم بیولوژیکی باشد که O 2 در آن پذیرفته شود ، می تواند یک ماده فعال ترش باشد. با این حال ، هیچ شرابخانه ای در واکنشهای آنزیمی تیره به دلیل وجود O2 وجود ندارد.
مدتهاست مشاهده شده است که سمیت نوری ترش مولکولی برای موجودات زنده در حال افزایش است. بسیاری از مردم از حضور در اتاق استقبال می کنند ، زیرا نور مرئی را لعاب می دهند - حساس کننده های نور 63 رنگدانه های طبیعی باگاتو می توانند حساس کننده نور باشند. در سلولهای موجودات زنده فتوسنتزی ، آنها دارای حساسیت زا فعال مانند کلروفیل و فیکوبیل پروتئین هستند. اکسیداسیون مولکولهای مهم بیولوژیکی تحت تأثیر نور مرئی در حضور اسید مولکولی و حساس کننده به نور ، اثر فتودینامیکی نامیده می شود.
63 حسگر نوری مولکول هایی هستند که می توانند نور و واکنش های شیمیایی را ایجاد کنند ، که در روز قابل مشاهده نیست. سبکی نور در مولکولهای کروموفورها ریخته می شود ، به طوری که می توان از هسته های چرخه ای انتقام گرفت. بیش از 400 کلمه وجود دارد که قدرت حساس کننده های نور هستند. در میان رودهای طبیعی ، حساس کننده های نور - کلروفیل ، فیکوبیلین ، پورفیرین و محصولات صنعتی ، سنتز ، تعدادی آنتی بیوتیک ، کینین ، ریبوفلاوین و غیره وجود دارد. حساس کننده های دایاک فقط در حضور O2 عمل می کنند ، اثر فتودینامیکی فعال می شود.
نور مرئی باید قبل از انتقال مولکول حساس کننده نور به اردوگاه آسیب دیده (* D) ریخته شود:
مولکول هایی که به اردوگاه مجرد منتقل شده اند ، می توانند در بوی بد فعالیت PHOTODYNAMIC به اصلی (D) تبدیل شوند یا به اردوگاه سه گانه هنوز زنده (* D) بروند. یک برچسب مکانیسم نصب شده است که در پشت آن می توان یک مولکول (* D) را به مولکول اکسیده شده به لایه اضافه کرد. یکی از آنها پانسمان پانسمان های تکی است. مولکول حساس کننده به نور در سلول سه قلو با O2 واکنش داده و آن را به یک اردوگاه جداگانه تبدیل می کند:
T D + O 2 D + * O 2.
بوسیله تک مولکول بستر (B) را اکسید می کند:
B + * O 2 BO 2.
اثر فوتودینامیکی تظاهرات در همه موجودات زنده پروکاریوتها ، در نتیجه عملکرد فوتودینامیکی ، تشویق می شوند تا انواع مختلفی را توسعه دهند: از دست دادن ظرفیت به شکل مستعمره ، از دست دادن DNA ، پروتئین ها و غشای سلولی. دلیل این امر ، پراکسیداسیون برخی اسیدهای آمینه (متیونین ، هیستیدین ، تریپتوفان و غیره) ، نوکلئوزیدها ، لیپیدها ، پلی ساکاریدها و سایر اجزای سلولی است.
مشتریان از سخنرانی ها ، نحوه تشخیص عملکرد خاموش کردن موسیقی منفرد و کاهش قدرت ساختاری و سایر انواع گفتار ، نحوه مشارکت انتقام می گیرند. یکی از "دودهای" ترش یکتا ، کاروتن است که موجودات فتوسنتز کننده را از اثرات کشنده ، حساس به نور به کلروفیل ، جذب می کند. پرهوپلاستی * O 2 є همچنین اسپولدهای فعال بیولوژیکی ایجاد می کند: لیپیدها ، اسیدهای آمینه ، نوکلئوتیدها ، توکوفرول ها و داخل.
ازن و بوسه اتمی.محصولات اسید مولکولی نیز اکسیژن اتمی (O) و ازن (O3) هستند. به نظر می رسد ، موسلین مولکولی در منطقه UV دور (160-240 نانومتر) به شدت رسوب داده است. بحث جداسازی فوتون ویکلیک مولکول به دو اتم:
O 2 + h 2O.
سپس خود به خود عکس العمل متقابل ، که منجر به تایید مولکول ازن می شود:
ازن می تواند در ترشح مولکولی در مشروب در طول تخلیه های قوی الکتریکی ، و همچنین در جریان آب الکتریکی و در برخی واکنشها ، در برخی موارد ، نظارت بر اکسیژناسیون ایجاد شود. اوزون و اسید اتمی اکسید کننده یاک قوی تر از O2 است. ازن را می توان با استفاده از انواع رادیکال ها عملا واکنش داد.
مقابله با مکانیسم های کلیتینی
برای خنثی سازی اشکال سمی گلبول های قرمز O2 ، آنها از مکانیسم های مختلفی استفاده می کنند که می توان آنها را به چند نوع تقسیم کرد. اساس سیستم ها برای نوع اول ، فعالیت آنزیم های ویژه ، برای توزیع اشکال سمی O2 ، اصلی و در تعدادی از انواع در یک عملکرد واحد است. در سیستم های مختلف ، برای از بین بردن اشکال سمی O2 ، تشکیل متابولیت های سلولی رخ می دهد. به عنوان یک قاعده ، سرنوشت سلولها در بازیابی سلولها از اثرات سمی O2 قدیمی تنها عملکرد متابولیتهای آنها نیست. نارشتی ، تا مکانیسم های خاص یک نوع خاص ، می تواند توسط تعدادی از پریستوسوان ها ، ریشه دار پروکاریوت ها در ریونیا قدیمی معرفی شود: جمعیتی ، فیزیولوژیکی ، ساختاری. عالی است ، اما بوی بد ساقه ها برای اهداف آنها ، دارچین و سم زدایی O2 است.
سیستم های آنزیمینسل اصلی انواع سمی آنزیمهای منسوخ O2 super: سوپراکسید دیسموتاز ، که مولکولهای O2 (واکنش 4) ، کاتالاز و پراکسیداز را مصرف می کند ، که H2O2 را جذب می کند:
برای تولید به حداقل غلظت در سلولهای O 2 -. і H 2 O 2 і به او توانایی همکاری با اظهارات HE را ندهید. (واکنش 5).
سوپراکسید دیسموتاز در پروکروتهای شیمیوتروفیک ، و همچنین O2 (شکلهای هوازی استان و اختیاری) ، و همچنین در نمایندگان زنده گروههای پروکاروتهای فتوسنتزی یافت می شود. در وسط آنزیم بی هوازی دانش در تعداد زیادی از اشکال هوازدگی. وینات قرار است دایاک باکتری های اسید لاکتیک شود ، پروتئین موجود در سلولها زیاد است ، آنها با غلظتهای بالا (حداکثر 30 میلی مولار) یونهای منگنز دو ظرفیتی جایگزین می شوند. وقتی صحبت از Mn 2+ می شود ، که برای آن نشان داده شده است حالت اکسیداسیون در حضور O 2 نشان داده می شود ، در چنین غلظتی از ساختمان ، تمیز کردن سوپر اکسید بسیار کارآمد است ، زیرا ممکن است سوپراکسید دیسموتاز را از بین ببرد ، سلولهای داخل سلولها را پاک کنید تا پلک بزنند. بنابراین ، در باکتریهای cich acid lactic ، عملکرد خنثی سازی O2 را Mn2+ می نامند.
در سلول های برخی از گونه های باکتری اسید لاکتیک ، هیچ سوپراکسید دیسموتاز یا غلظت بالای Mn2+ یافت نشد. Tsi vidi با حساسیت بیشتری نسبت به O2 مشخص می شود.
در میان بی هوازی های منطقه ای ، سوپراکسید دیسموتاز در نمایندگان باگاتوخ جنس یافت می شود کلستریدیوم... Vivchennya їkh stikosti تا O 2 vyavlya یک پیوند واضح در محل آنزیم در سلول ها است. می بینید که سوپراکسید دیسموتاز بسته به گونه ای که آنزیم در آن فعال است ، می تواند با قدرت نسبتاً بالایی تا O2 مشخص شود. سوپراکسید دیسموتاز در سایر گونه های باکتری های کاملا بی هوازی شناخته شده است. تعداد موجوداتی که هنوز با سوپراکسید دیسموتاز کشف نشده اند هنوز اندک است.
ظاهر شدن سوپراکسید دیسموتاز در بی هوازی های شدید (وسیع تر ، کمتر در اوایل مجاز) تغذیه را در مورد نقش فیزیولوژیکی موجودات زنده متمایز می کند. سلامت بقیه رشد تنها در محیط عاری از اسید است تا عملکرد نامعقول آنزیم را در مغزهای داده از بین ببرد. شما می توانید ، تنها در صورتی که برای یک هوازی جدید بی هوازی ناخوشایند انجام دهید ، سنتز آنزیم توسط اسید مولکولی فعال می شود ، که از جذب O2 در ذهن در گردش جلوگیری می کند.
سوپراکسید دیسموتاز آنزیمی است که می تواند در مرکز فعال گروه پروتز و فلز انتقام بگیرد. در پروکروتها - اتمهای منگنز و / یا ناهنجاریها. تعداد زیادی از سوپراکسید دیسموتاز توسط دو زیر واحد یکسان ایجاد می شود که پوست آنها انتقام یک اتم فلز را می گیرد. آنزیمهای Fe- و Mn برای عوارض بعد از اسید آمینه انتخاب می شوند. در تلاش برای یافتن ارتباط بین ویژگیهای فیزیولوژیکی و سایر ویژگیهای موجودات و اشکال فلزی ، آنزیم منجر به تغییر کافی در آنها نشد. شکل اول و دوم سوپراکسید دیسموتاز در نمایندگان پروکاریوتهای گرم مثبت ، عکسهای میانی کموتروفها ، بی هوازیهای منطقه ای ، هوازیها و اشکال اختیاری بی هوازی یافت شد. علاوه بر این ، متالوفرم های تهاجمی سوپراکسید دیسموتاز می توانند در یک ارگانیسم وجود داشته باشند و وارد مولکول یک آنزیم شوند. برای برخی از گونه ها ، نشان داده شده است که سنتز همان نوع آنزیم در حضور یون های فلزی در وسط کشت قرار دارد.
سوپراکسید دیسموتاز در پروکاروتیدهای کموتروفیک داخل بدن آنزیمی است که به غشاها متصل نیست و در سیتوپلاسم قرار دارد. دارند E. coliدر سلولهایی که شکل Fe- ، Mn- و Fe / Mn به آنزیم منتقل شده است ، Fe- سوپراکسید دیسموتاز در پری پلاسم و Mn آنزیم را در سیتوپلاسم قرار می دهد. در اتصال سایم ، خورشتی وجود دارد که آنزیم متالوفرم در تشکیل سلولها به شکل O2 نقش دارد -. : آنزیم را از آنیونهای سوپراکسید برونزا و Mn-yaku-از آنیونهای درون زا بگیرید.
به خصوص مشکل خلاص شدن از شر پیری و مولکولی در سلولهای سیانوباکتری یک مشکل بسیار جدی است. Ymovіrno ، بوی بد اولین در بزرگترین جهان ، وراثت اثرات سمی ترش را مشاهده کرد. سوپراکسید دیسموتاز در همه سیانوباکتری ها یافت می شود. در کلیتین ها Anacystis nidulans (Synechococcus) Fe-superoxide dismutase ، که تا 90 of منشاء آنزیم ذخیره می شود ، در سیتوپلاسم کلتین قرار دارد و شکل Mn-yaka در Tylakoid است. عملکرد آخرین شکل آنزیم بلافاصله تا زمان اضافه بار یون O2 -تولید می شود. ، در فرآیند انتقال الکترونیکی فتوسنتز شرکت می کنند.
کاتالاز و پراکسیداز.پراکسید در دو دسته آنزیم های اسپور از بین می رود ، که تبدیل دو الکترون به H2O و ویکوریست به عنوان اهدا کننده الکترون H2O2 در مورد کاتالاز (واکنش 10) یک ارگان را کاتالیز می کند.
فعالیت کاتالاز و پراکسیداز در همه پروکروتهای هوازی اجباری و اختیاری رخ می دهد. در میان بی هوازی ها و آنزیم های منطقه ای در جهان کوچکتر ، سوپراکسید دیسموتاز کمتر گسترش می یابد. بسیاری از بی هوازی های سخت و هوازدگی آشکار شده است ، که عبارت است از انتقام از سوپراکسید دیسموتاز ، یا انتقام نگرفتن از کاتالاز. تا تاریخ چهارم ، ممکن است آن باکتریهای اسید لاکتیک را وارد کنید ، که در آنها یونهای O2 ایجاد می شود -. مراقب Mn 2+ باشید که در سلولهای نزدیک غلظت بالا یافت می شود.
شیوع کاتالاز در باکتری های اسید لاکتیک با آن ارتباطی ندارد ، زیرا بوی بد نمی تواند هم را سنتز کند - یک گروه مصنوعی از آنزیم ، که برای سنتز آپوآنزیم کافی نیست. با افزودن گروه های هم ، باکتری های اسید لاکتیک کاتالاز حاوی هم را تشکیل می دهند. در تعدادی از باکتری های اسید لاکتیک ، کاتالاز یافت می شود که از گروه خونساز انتقام نمی گیرد ، به آن کاذب نامیده می شود. آنزیم بینایی در شش قند پلیپپتیدی یکسان ، با نیروهای غیر کووالان ذخیره می شود. زیر واحد پوست برای انتقام 1 اتم منگنز.
آب پراکسید ، که نتیجه اثر متقابل ویتامین O 2 است ، توسط مسیرهای غیر آنزیمی جذب می شود. به نظر می رسد Fe 2+ در محلول آب باعث ارتقاء H 2 O 2 به H 2 O می شود. مقدار کمی یون در سلول ها وجود خواهد داشت. Ruinuvannya H 2 O 2 را می توان برای راخونوک مشاهده کرد که در وسط فرهنگی بومیان محلی تجدید شده دیده می شود.
برای پروکاریوتهای بی هوازی ، که می توانند تماس با O2 را تحمل کنند و هنگامی که آنها در مقیاس کوچک وجود داشته باشند ، وجود سوپراکسید دیسموتاز در سلولها ضروری است ، "تمیز کردن" O2 -. ... وجود کاتالازی ها ، در صورت لزوم ، برخی از مجاورت ها در واکنش تغییر شکل و در واکنشهای تجزیه پراکسید به صورت خود به خود ، برای مشارکت کاتالیزورهای غیر آنزیمی ، و موجودات در کل نمی توانند با آن کنار بیایند. بنابراین ، وقتی متابولیسم پرانرژی است ، برای کاهش اثرات سمی O2 از نوع بی هوازی است که به اندازه کافی یک غشای آنزیمی در سوپراکسید دیسموتاز وجود دارد.
افزایش مقیاس تعامل پروکاروئیدها با O2 در صورت عملکرد متابولیسم هوازی ، برای جلوگیری از روشهای غیر موثر آنزیمی استفاده از H2O2. کاتالاز و پراکسیداز دریافت نکنید. در چنین رتبه ای ، در ذهن فعل و انفعال فعال بین سلولهای O 2 ، به طوری که بتوانیم هر نوع زندگی هوازی را از بین ببریم ، سیستم جذب آنزیمی اثرات سمی با مشارکت سوپراکسید دیسموتاز ، کاتالاز و پراکسیداز در اجزای لازم (شکل 87).
مکانیسم های کمک گرفتن از متابولیت های سلولی Zachist در برابر یکی از سمی ترین O2 قدیمی - ترش منفرد - به کمک مولکولهای جدید بیولوژیکی مهم بروید. همه انواع خاموش کننده * O 2 را می توان به فیزیکی و شیمیایی تقسیم کرد. آنها آن را فیزیکی می نامند ، زیرا منجر به خرابی گازنیک ها نمی شود:
* O 2 + A O 2 + A
تولید گاز تا زمان اکسید شدن بسیار مشکل است:
* O 2 + A O 2 + A خوب
اشباع شدن مکانیسم شیمیایی خاموش کردن O2 با اسیدهای چرب ، لیپیدها ، اسیدهای آمینه ، نوکلئوتیدها و سایر قطعات مهم است. مکانیسم های خاموش کننده شیمیایی تکاملی ، البته در انواع زیادی از مرحله بلال ، ایجاد پراکسید چرخه ای لب با توزیع های بیشتر ، گویی تغییرات بنیادی را ایجاد می کند. شیمیایی خاموش شده * O2 را می توان در سلول ها تا سوته وراثت مخرب آنها تولید کرد. قبل از خاموش کردن مکانیسم اصلی فیزیکی ساخت مولکول های دیگر spoluks شیمیایی. م Mostثرتر در انواع کاروتنوئیدها ، که به طور گسترده در باکتری های سبک گسترش یافته است. بوها در سلولهای کموتروفهای هوازی باگاتوک ، یک جزء لزج کلی از دستگاه رنگدانه همه فوتوتروفها یافت می شوند. در سلولهای موجودات فتوسنتزی ، یاک به عنوان حساس کننده فعال فعال ، کلروفیل تعیین می شود. با این حال ، احتمال اثرات اکسید کننده عکس در ذهن عملکرد دستگاه فتوسنتز می تواند از طریق یک ساعت فوق العاده کوتاه (10 تا 11 ثانیه) کلروفیل در یک سلول تحریک شده به سطح پایینی برسد و به طریقی دیگر ، از طریق فوتوکسیداسیون یک سلول سرطان زا.
برای اولین بار ، نقش کاروتنوئیدها در اثر کشنده ، مانند فوتوکسیداسیون ، در مورد تلقیح جهش یافته بدون کاروتنوئید باکتری بنفش نشان داده شده است. Rhodopseudomonas sphaeroides... فرهنگ خوب در ذهن های مختلف از نظر فوتوتروفی رشد می کند ، اما می تواند در نورها و در خلق و خوی در ذهن های خارجی رشد کند. حذف جهش یافته از او ، افزودن کاروتنوئیدها ، رشد ناچیز نور در ذهن های مختلف و خلق و خوی در ذهن های هوازی ، هنگام انتقال به نور + وحی کاملا خجالتی است.
فرآیندهای اکسیداسیون نوری می توانند در پروکاریوتهای غیر فتوسنتز و همچنین مولکولهای موجود در سلولهای آنها ایجاد شوند که نور مرئی را لعاب می دهند ، زیرا می توانند به عنوان حساس کننده نور عمل کنند. دیه کاروتنوئیدها تنها با مشارکت آنها در فرایند یک اثر فتودینامیکی در هم تنیده نیست. بوی تعفن اردوی تک نفره به دلیل این واقعیت است که در برخی واکنش های برندگان: روی چراغ ها یا در temryavi ، بوی بد ترش می شود.
مکانیسم توسعه کاروتنوئیدها در ارگانیسم های فتوسنتز کننده میدان در حمله (شکل 88). مولکول کلروفیل ، که نور ریخته است ، به سرعت (10 -12 ثانیه) انرژی انرژی منفرد را به مرکز واکنش منتقل می کند. حدود 104 کوانتوم نوری برای تولید تقریبا 4 قبل از انتقال مولکول کلروفیل به آسیاب سه قلو آسیب دیده تولید می شود. به دست آوردن قدرت ضربه فوتودینامیکی. کاروتنوئیدها می توانند در سه واکنش شرکت کنند: 1) بدون نیاز به خاموش کردن سه گانه کلروفیل و انتقال آن به طبقه اصلی (شکل 88 ، آ)؛ هنگامی که مولکول سه گانه کاروتنوئید باعث ایجاد انرژی اضافی در گرما می شود و به اردوگاه اصلی تبدیل می شود. 2) کلروفیل سه گانه توسط کاروتنوئیدها خاموش نمی شود. vidbuvatsya yogo vzaєmodiya z O 2 ، scho ترجمه ostnіy به اردوگاه تک نفره porusheny ؛ بوسه منفرد توسط کاروتنوئیدها خاموش می شود (شکل 88 ، ب)؛ 3) تک بوسه ، که توسط کاروتنوئیدها از مکانیسم فیزیکی خاموش نشده است ، می تواند با آنها در ارتباط باشد واکنش های شیمیایی، Scho منجر به اکسیداسیون کاروتنوئیدها می شود. سرنوشت کاروتنوئیدها در هر یک از سه واکنش ذکر شده در بالا ، سطح آگاهی سلول ها را کاهش می دهد * O2.
وجود پروکاریوتها ، که به آنها در بسیاری از انواع اثرات سمی اسیدی شدن مولکولی کمک می کند.در سلولهای کلستریدیوم بی هوازی اجباری بی سر و صدا ، که در آنها سوپراکسید دیسموتاز ، کاتالاز وجود ندارد ، از راهی برای خنثی سازی O2 برای افزایش کارایی کشت استفاده می شود ، اما در تامین گاز نیز فعال است (CO 2) تعلیق بحرانی ترش از وسط ، که منجر به خم شدن قسمتی از کراتین می شود ، اگرچه ممکن است بیش از حد در سینک های کاهش یافته O 2 تکثیر شود.
با باکتری های اسید لاکتیک در یک خط مستقیم از ترش مولکولی تکه تکه شدن ، کروکوس آواز خواندن به جلو. Cybacteria یک گروه از پروکاریوت ها هستند که فاقد کاتالازهای حاوی هم هستند که در حضور یک نوشیدنی رشد می کنند. سر و صدای مکانیسم های خنثی سازی O2 و افراد مسن منجر به ایجاد سوپراکسید دیسموتاز و غلظت داخلی بالای یونهای Mn 2+ شد ، از زمان انتشار O2 -. ، سودوکاتالاز ، و همچنین مانند فعالیت کاتالاز و پراکسیداز. در برخی از نمایندگان باکتری های اسید لاکتیک ، می توانید خمیدگی واضحی از پاهای چسبیده به O2 را مشاهده کنید ، که می تواند برای تولید دارچین خوش طعم ویکوریستان استفاده شود. برای کودکان باکتری اسید لاکتیک از جنس لاکتوباسیلوستوزیع سریع گلیکولیتیک گلوکز در سینک های هوازی را نشان می دهد. با این موضوع ارتباط دارد که در غرق های هوازی ، آب NAD-H2 می تواند مستقیماً به O2 ، بخشی از اسید پیروویک از عملکرد گیرنده منتقل شود ، زیرا در اثر تخمیر اسید لاکتیک سرپانتین ایجاد می شود. اسید پیروویک اکنون می تواند از زنجیره "اتصال" به استیل-کوآ اکسید شده و سپس برای تولید سنتز یک مولکول ATP به استات متابولیزه شود. ممکن است bachiti ، سرنوشت ترش شدن در کل فرایند به طور مستقیم با رد انرژی روحانی مرتبط نیست (هنگام انتقال آب از NAD-H 2 به O 2 ، انرژی به شکل ATP انجام نمی شود) ، به عنوان مثال O 2 ، یک وظیفه پذیرنده ، بخشی از پیروات را بر عهده بگیرید ، همانطور که می توانید از مسیر پرانرژی vikoristovuyuyu کنید ، اما در نهایت کیسه ای را برای خروجی پر انرژی سرگردان نشان دهید. بنابراین ، اکسیداسیون مستقیم بخشی از حامل های به روز شده الکترون در روند سرگردان ممکن است نه تنها منفی ، بلکه وراثت مثبت باشد.
Yak به معنی ویل ، حتی حساس به فرآیند تثبیت نیتروژن O2 بود. ساختمان فیکسوواتی شماره 2 بدون دخالت در قیمت وسط پروکاریوت ها به طور گسترده ای گسترش می یابد ، بنابراین ترش مولکولی افزایش می یابد. قدرت شیمیوتروفها و فوتوتروفها ، از جمله سیانوباکتریها ، مسئول فتوسنتز اسیدی است. Fiksuvati N 2 می تواند اشکال آزاد و پروکاروتیدها باشد که در همزیستی با موجودات یوکاریوتی یافت می شود.
دخالت در روند تخریب در پروکاریوت ها نشان داد که در بسیاری از موارد ، از کارایی 100٪ دور است. در بین تثبیت کننده های نیتروژن هوازی می توان شاهد محرومیت از موجودات زنده ، رشد در وسط N2 در ذهن کودکان بود. احتمال і fiksuvati N 2 فقط در ذهن کاهش غلظت ترش مولکولی ، به عنوان مثال ، در ذهن میکرو ایروبیک بیشتر است. نیتروژناز زاکیستی در سلولهای بی هوازی اختیاری کمتر م :ثر است: بوی بد می تواند نیتروژن را فقط در ذهن بی هوازی ثابت کند.
نمایندگان این جنس با تعداد ثابت کننده های نیتروژن هوازی آشنا می شوند. آزوتوباکتر ،در کسانی که در تصرف آنها ظاهر شده است. یکی از آنها با بهبود شدید فعالیت غشایی سلول ها ، که مسئول تثبیت نیتروژن در مغز هستند ، مرتبط است. انرژی به معنای جهان این است که به طرق مختلف برای "اتصال" غشای داخلی O 2 عمل کند ... دیچنا لانتسیوگ Azotobacter vinelandiiبرای به پایان رساندن توزیع آسان و تاشو در جاده ها تا انتقال الکترون ها به سیتوکروم بحمل و نقل الکترونها ، پانسمان از فسفوریلاتها ، حرکت از طریق:
سیتوکروم ب ج 4 ج 5 آ 1 .
هنگامی که یک وضعیت "سرگیجه" وجود دارد ، رشد فعالیت حمل و نقل الکترونیکی بر روی دستها: سیتوکروم ب دبه ذخیره انرژی وابسته نیست. Tse تا حدی تولید شود که بدون تأثیر بر رشد فعالیت خارجی ، تأثیر به دست آوردن حمل و نقل الکترونیکی با ذخیره انرژی کاهش می یابد. در چنین رتبه ای ، "سوزاندن" بخشی از بسترهای کربنی در نظر گرفته می شود ، زیرا می توان از آن برای تجدید O2 بدون ذخیره مقدار زیادی انرژی استفاده کرد.
علاوه بر "قربانی شدن" ، قسمتهایی از dzherel در غلظت بالای O2 به سلولهای گرگینه ساختار نیتروژناز بد می آید ، به طوری که حساسیت به ترش مولکولی ظرف برای هر کسی کمتر قابل دسترسی است. Vislovlyuyuyutsya rіznі pisushennya علاوه بر این ، به عنوان یک "سازگار" zahist وجود دارد. شما می توانید ، در همان زمان ، تغییر در رشد متقابل دو نیتروژناز ایجاد شود. سرنوشت این نوع غشای سلولی گنجانده نشده است. آواز تثبیت نیتروژناز در مغز با غلظت بالای O2 به مجموعه آنزیمی کاتیونهای دو ظرفیتی اضافه می شود. نارشتی ، سطل های مخصوص ظهور ، که مجتمع هایی با سطل های نیتروژناز ایجاد کرده و در حضور O 2 پایداری ایجاد می کند.
مقدار زیادی پروکاروت حاوی نیتروژن در تأمین نیتروژن مولکولی در فاضلاب های ریز. تا زمان پیوستن ژیسنیخ به آنها ، آنها عبارتند از: ایجاد لیس ، که انتشار را به سلول O 2 منتقل می کند و به خودی خود یک ناحیه میکرو هوازی را در نزدیکی آن باز می کند. شکل گیری خرید مشتریان ، سهولت دسترسی به O 2 به مشتریان ، roztasovanim وسط خرید ، که در چنین رتبه ای ، برای تولید نیتروژن دوستانه تر است. تشخیص گونه های تثبیت کننده نیتروژن در ارتباط با هتروتروفهای هوازی غیر تثبیت کننده نیتروژن ، که گونه های تثبیت کننده نیتروژناز نیتروژناز را از دسترسی O2 به دست می آورد.
ضمیمه ویژه برای جذب نیتروژناز در غلظت های بالای viroblency O2 توسط ثابت کننده های نیتروژن همزیستی - باکتری های پیازی. همراه با خود پیازها ، روند انتشار فعال باکتری ها و تثبیت آنها N 2 ، پس از مشاهده ساختار ، یکی از نشانه های آن اتصال متقابل دسترسی به وسط اسیدیته مولکولی است. وظیفه ویکون این است که از پیازهای لگهموگلوبین (بلوک مشابه هموگلوبین) انتقام بگیرد ، که به طور فعال O2 را می خواند و عملکرد مناسب آن را در باکتری کنترل می کند. هرکسی که متابولیسم سالمی از نوع هوازی انرژی داشته باشد ، تحت اسیدیته مولکولی انباشته قرار می گیرد.
Naybіlsh gstroit یک مشکل برای فرآیند تثبیت نیتروژن O2 در گروه سیانوباکتری ها است. در همه سیانوباکتری ها ، نیتروژناز به O2 حساس است و هیچ فعالیت خارج سلولی و درون سلولی وجود ندارد. در حقیقت ، آنها می توانند مشغله ای را ببینند که آنها را به سمت اسیدیته برون زا سوق می دهد و برای خنثی سازی O2 در نظر گرفته شده اند تا در فرایند فتوسنتز خود را در وسط سلولها مستقر کنند.
اگر همه سیانوباکتریهای تثبیت کننده نیتروژن از پایین مرحله تا فرآیند تثبیت نیتروژن از O2 دیده شوند ، می توان آنها را به دو گروه تقسیم کرد. قبل از گروه اول ، سیانوباکتریها وجود دارند ، در برخی از آنها تثبیت نیتروژن O2 م effectiveثرترین است ، بنابراین سلولهای رویشی می توانند N2 را فقط در نوعی میکرو هوازی انجام دهند. گروه دیگر دارای سیانوباکتری ها هستند ، که برای تثبیت نیتروژن سالم در هوا شستشو می شود ، سلولهای خاصی تشکیل می شود - هتروسیست.
در سیانوباکتریهای غیر هتروسیتیک ، فعالیت سیانوباکتریوم سیانوژناز فعالیت سیانوژناز رویشی از O2 است ، در وهله اول سیانوباکتریهای درون زا ، در فرآیندهای فتوسنتز و فعالیت نیتروژناز ، بدون وقفه در سنتز ، پشتیبانی اضافی وجود دارد. از اکسیژن رسانی در مرکز رشته های برخی از اشکال غیر هتروسیتی ، سلولهای رویشی با رنگدانه ضعیف اغلب مشاهده می شوند ، در برخی از آنها اغلب ظرفیت فتوسنتز CO2 دیده می شود. (این هتروسیستها نیست ، ala ، imovirno ، از آنها هتروسیستها مرکز جک تثبیت نیتروژن را در سینک های هوازی ایجاد کرده اند). به منظور سنتز O2 برون زا ، می توان از سنتز مقدار زیادی مخاط ، اغلب کلیتین ناولکولیشی از سیانوباکتری های نیتروژن دار استفاده کرد. آگاهی از دیدگاه اشکال استعماری همچنین می تواند از ظهور ذهن های بی هوازی برای کلیتین هایی که در قسمت مرکزی مستعمره بزرگ شده اند جلوگیری کند.
نیبیلش اسیدی شدن مولکولی درون زا و برون زا را در هتروسیست ها به طور کامل مطالعه کرده است. هتروسیستها تا حد فتوسنتز O2 رشد نمی کنند. اما فعالیت زیاد مخلوط اکسید پنتوز فسفات ، که برق را به لنز دیکلروم تأمین می کند ، توسط O2 پذیرفته نمی شود ، سوپراکسید دیسموتاز را آزاد می کند تا لخته شدن روغن گیاهی آزاد شود. bar'єru ، - تمام راه برای جذب سیستم نیتروژنی در هتروسیستها به عنوان فعال سازی توسط اسید مولکولی.
در چنین رتبه ای ، می توان فقط مجاز بود ، اما با مکانیسم های خنثی سازی ترشح مولکولی در مراحل جدید تکامل همراه با آن ، کلیتین بولی یکسان نیست. H2 مرحله ای از واکنش های آنزیمی است که گنجاندن O2 در متابولیسم پروکاروت ها را کاتالیز می کند.
اکسیژن مولکولی در متابولیسم پروکاروئیدها
واقعیت این است که همه کسانی که روی زمین پروکاریوتی هستند ، بی هوازی های گوشتی را می بینند ، در حضور O 2 ، آنها را لعاب می دهند ، تا بر واکنش های اسیدیته مولکولی به آنها تأثیر بگذارند. با توجه به نسبت تا O2 ، همه زایمان ها را می توان به چند گروه فیزیولوژیکی تقسیم کرد (شکل 34). چنین پیدروزدیل در مورد نیاز به مکتب ترش مولکولی صحبت می کند ، نه اینکه مکانیسم های تعامل با او را باز کند. Dіysnі ، بلافاصله می دانیم که O2 می تواند کلید ضروری برای حذف انرژی یا به خاطر همه واکنش های مشابه باشد ، زیرا ارزش انرژی ندارد.
کم اهمیت. 89. Wickers victoriannya portsii ترش مولکولی ترشح شده با گوه. برای توضیح ، متن را ببینید (بعد از اسکولاچوا ، 1969).
H. P. Skulachov چنین طبقه بندی واکنش هایی را در تعامل سلولین با اسید مولکولی پیشنهاد کرد (شکل 89). بخشی از سلول O2 رسوب شده را می توان به دو قسمت مختلف تقسیم کرد. توده اصلی غذای ترش از سلولها برای مشارکت سیستمهای آنزیمی سلولی تشکیل شده است. رسوب بخش سلولز O2 به سیستم آنزیمی وابسته نیست. استراحت Ilyustratsієyu برای خدمت به خوبی از تمیز کردن فعال دوغاب کشته شده با گرم کردن clitins. در همه انواع ویپادکا ، تعقیب و گریز صرفاً انجام می شود روند شادی، پانسمان با شیمی آواز سلولی اکسید شده ، به عنوان مثال ، گروههای SH سلولهای سلولی. غیرممکن است که امکان اجرای فرایندهای مشابه در تعلیق سلولهای زنده وجود داشته باشد. با هزینه خود ، ترشح مولکولی آنزیمی - انرژی 64 - برای اکسیداسیون رشد می کند ، اما نه برای ذخیره انرژی و همچنین برای ذخیره انرژی ، یعنی با ذخیره انرژی برای سلول ارتباطی ندارد. واکنشهای آنزیمی اکسیداسیون برای مشارکت O2 ، که می توانند به دسته اکسیداسیون حیاتی منتقل شوند ، واکنشهایی هستند که در نتیجه آنها می توان انرژی را در گرما 65 مشاهده کرد. قبل از کل دسته بندی فرایندها ، واکنشها انجام می شود ، که شامل تک اکسیداسیون است. به مولکول گفتار قابل اکسیداسیون ، و همچنین واکنش هایی که برخی از اکسیدازها را کاتالیز می کند.
اصطلاح "dichannya" برای اولین بار مقدماتی را برای روند معنی دار آواز ، مرتبط با زندگی سایر موجودات زنده (رزلین و توارین) معرفی کرد. دو ویژگی اصلی این فرایند را مشخص می کند: تبادل گاز از سطح میانی ثابت با مشارکت نابرابر O2. نیاز به زندگی بدن. شباهت اصلی به فرآیند دوگانگی در سطح روحانی در همه موجودات دیگر ، کاشت یک اصطلاح را به روشی آسان از بین برد و فهمید که این درک قابل خواندن است. برندگان تاشو هنگامی که اصطلاح "dichannya" به دلیل تطبیق پذیری فوق العاده برای معنی فرآیندهای عملکردی مشابه در پروکاریوت ها راکد است. در ذهن ما ، اصطلاح "dichannya" در تمام فرآیندهای رسوب آنزیمی با یک سلول ترش مولکولی گسترش می یابد.
واکنشهای اکسیداسیون طبیعی ممکن است در تنظیم حرارتی سالم در حیوانات در طول خنک کننده موجودات مهمتر باشد.
تجمع آنزیمی O2 ، به دلیل ذخیره انرژی ، برای فرایندهایی که با فسفوریلات ارتباط ندارند ، و فرآیندهایی که روی فسفوریلات ها چیده می شود ، مورد استفاده قرار می گیرد. اولین نوع اکسیداسیون ، به دلیل ذخیره انرژی ، به تبدیل انرژی به شکل پیوندهای فسفاته کلان وابسته نیست. ظاهراً دو شکل کلی انرژی در اتاق وجود دارد: شیمیایی و الکتریکی ( H +). یکی از راههای رد انرژی به شکل شیب الکتروشیمیایی غشایی H + با انتقال الکترون به O2 پیوند می خورد. انرژی شیمی عمدتا در زیرزمین ذخیره می شود ، جایی که پیوندهای فسفات ماکروارژیک را جایگزین می کند ، و در محل قرارگیری ، در مولکول های ATP ذخیره می شود. در مراحل میانی فرایندهای کاتابولیک ، در کیسه انتهایی با رسهای O2 گره خورده است ، متابولیسم ایجاد می شود ، که پیوند انرژی زیادی را نشان می دهد ، به عنوان مثال ، انرژی (C ~ S - CoA). Tsi spoluks می تواند بدون پیش فرض zaprepuvaty energієyu deyakі فرآیندهای بیوسنتزی.
در همان زمان ، هنگامی که انرژی اکسید شده فسفوریله می شود ، در حین انتقال الکترونیکی به صدف مولکول و انگور به شکل +H +، پس از سنتاز ATP پروتون اضافی تبدیل می شود تا به شکل شیمیایی در مولکول های ATP تبدیل شود. بر اساس همه موجودات دیگر ، سطح بالایی از انتقال اسید فسفریک به دست می آید ، به عنوان مثال E. Tsei shlyakh در حال حاضر در نمای تشکیل شده است ، در سایر پروکاریوت ها ، انتقال الکترون ها و فسفریلات ها منتقل می شود. همه انواع فرآیندهای اکسیداتیو به دلیل مشارکت O2 ، که در سلولهای بسیار سازمان یافته رخ می دهد ، در پروکاریوت ها ظاهر می شود.
اساس طبقه بندی ، که توسط V. P. Skulachov پیشنهاد شده است ، ردیابی تمام واکنش های موجود در رابطه بین سلولین و ترشح مولکولی انرژی است. برای مکانیسم های شیمیایی زیر این واکنش ها ، همه بوی بد را می توان به 2 نوع تقسیم کرد. قبل از نوع اول ، واکنشهایی انجام می شود که کاتالیز کننده ترانسفراز اسیدی یا دیوکسی ژناز است ، در برخی موارد ، مولکولهای اسید را مستقیماً به مولکولهای متابولیسم منتقل می کند:
A + O 2 AO 2.
یک مولکول به بستر می تواند جرم اتم مولکول و همچنین در جهت واکنش را بپذیرد. گیرنده های O2 می توانند مولکولهای دو لایه متفاوت باشند:
X + Y + O 2 XO + YO.
همه واکنشهای دیگر اکسید نشده و به نوع الکتریسیته مورد استفاده مربوط نمی شوند.
در واکنش انواع مختلف الکترونیک ، به تابع اسیدی بروید ، که عملکرد گیرنده انتهای جنس است. در همان زمان ، 1 ، 2 یا 4 الکترون در حضور طبیعت حامل توسط مولکول ترش پذیرفته می شود ، که می تواند در کیسه انتهایی به یک تصفیه نشده (O 2 ، H 2 O 2) یا اضافی تولید شود. (H 2 O) تجدید. واکنش های این نوع توسط آنزیم هایی به نام اکسیداز کاتالیز می شوند و می توانند بدون ذخیره انرژی ، بسیار اکسید و اکسید شوند. قبل از واکنشهای اکسیداسیون ویلنی ، واکنشهایی انجام می شود که توسط رزین اکسیدازها کاتالیز می شوند و در سیتوپلاسم سلولینی قرار دارند. به غیر از آنها ، در توصیف پروکاریوتها ، تعدادی از اکسیدازها که با غشاهای سیتوکروم و غیر سیتوکرومی مرتبط هستند ، انتقال الکترونها از آنها به O2 نیز با ذخیره انرژی حاصل نمی شود.
با واکنش های پشت مکانیسم شیمیایی و ایجاد واکنش های غذایی ، که در آن نسبت پوست دو اتم در مولکول اسیدی است:
A + O 2 + DH 2 AO + H 2 O + D
در وهله اول ، 1 اتم از مولکول تخریب شده به دلیل اتصال مستقیم به آن برای اکسیداسیون گفتار اسیدی می شود. و در صورت اهدا کننده مناسب لوازم الکترونیکی به H 2 O به روز می شود. جرم واکنشها توسط یک آنزیم کاتالیز می شود ، که می تواند منجر به گروهی از مونوکسیژنازها یا اکسیژنازها (اکسیدازها) با عملکردهای مختلف شود. مونوکسیژنازها در سلولهای جمعیت و تکامل. بوی بد واکنش اکسیداسیون را کاتالیز می کند. مشارکت در فرآیندهای مرتبط با ذخیره انرژی ، اندک است.
بنابراین ، اکسیژناز آنزیمی است که فعال شدن O2 را کاتالیز می کند و 1 یا 2 اتم را در مولکول های دیگر بسترها شامل می شود. بستر Yaksho (O2 گیرنده) آب است ، آنزیم اکسیداز نامیده می شود. در مفهوم وسیع اکسیداز ، می توان تخصصی از کلاسهای اکسیژناز را مشاهده کرد.
اکسیژنازها نقش مهمی در فرایندهای بیوسنتز ، تجزیه و تبدیل متابولیت های سلولی دارند: اسیدهای آمینه معطر ، لیپیدها ، آب نبات ، پورفیرین ، ویتامین ها. بسترهایی که اکسیژناز روی آنها تزریق می شود ، اغلب به عنوان یک نوآوری قوی در تامین آب عمل می کنند. برای تولید اکسیداسیون قبل از قوی شدن محصولات واکنش در آب و همچنین از نظر بیولوژیکی فعال ، که برای متابولیسم بیشتر مهم است. در پروکاریوتهای کاملا بی هوازی ، بوسه ، که در مولکول بستر موجود است ، از O2 جذب نمی شود ، بلکه از دیگران ، به عنوان مثال ، آب جذب می شود.
از همان نقطه ، کل تعامل منابع مولکولی با یک سلول ، از نظر قرار گرفتن در اساس مکانیسم های شیمیایی ، می تواند در مشارکت O2 در دو نوع واکنش ، در اولین واکنش که به شکل یک چرخنده در مولکول گفتار قرار می گیرد. تنها اولین نوع واکنش برای مشارکت در ترشح مولکولی می تواند به منبع انرژی سلول تبدیل شود. به همین دلیل است که برای ما مهم است که تحولات متقابل سلول ها با O2 را در مسیر شکل گیری سیستم هایی توسط آن ، که شامل ترش مولکولی در نقش گیرنده الکترونیکی است ، تحلیل کنیم.
فرم "مکانیزم اکسیداسیون" VZAЄMODIS Z kissen مولکولی ، موجود در انبار ENERGIS
با ظهور O 2 در جو ، امکان انتقال به لوازم الکترونیکی جدید. برای انتقال مقدار کمی پانسمان از بین بردن انرژی ، لازم است یک فانوس حمل و نقل الکتریکی با درجه آواز ، که توسط حامل ها در غشاها حمل می شود ، تدوین شود تا از انتقال پروتون ها به مرحله اول در دستگاه الکترونیکی دیگر جلوگیری شود. انتقال انرژی الکتریکی در شیمی ، که در مولکولهای ATP ذخیره می شود.
زی انجام elektrontransportnuyu lantsyugami، lokalіzovanimi در membranі scho در mіstyat OOO همه TIPI perenosnikіv من mayutsya مستقیم vіdnoshennya به otrimannya klіtinoyu energії، M- Vzhe zustrіchaєmosya در rozglyanutih در rozdіlah 13 14 eubakterіy معمولی بدن naybіlsh به سادگی organіzovanoyu انرژی chemotrophic (brodіnnya) من phototrophic (bezkisnevomu نوع فتوسنتز): همه باکتری های اسید پروپیونیک ، همه باکتری های بنفش و سبز فتوسنتزی. در غشای سلولی ، ارگانیسم های cich موضعی هستند و از نظر عملکردی با انتقال الکترونیکی سنتاز ATP ارتباط دارند.
P. Mitchell نشان داده است که سیستم انتقال الکترون ها و پروتون ها و انتقال پروتون های ATPase یکی یکی نصب می شود ، اما نه یک شبه به عنوان روشی متفاوت برای تولید H +، که برای یک فرایند ارتعاشی برای ایمن سازی اتصال ضروری است. به دلیل "دستگاه" هر دو سیستم ، گوش در نتیجه فرایندهای انتقال الکترون و فسفوریلاسیون در نتیجه تخریب ربات ATFazi در سلول ها باقی ماند. قیمت تجمع انرژی اکسید شده در مولکولهای ATP را کاهش داد. یک انبار نزدیک و ساختار مشابه غشاهای تبدیل کننده انرژی ، شباهت زیادی با مکانیسم های بدست آوردن گروههای بدون پروسوت بدون فسفر و مقرون به صرفه ، از جمله آنهایی که بر اساس مراحل اولیه تکامل ، سیستم بدست آوردن ارگانیسم الکترونیکی هستند
درباره راه رفتن در اطراف پروتون معکوس ATFazi
نیبیلش مدتهاست که یک ATPase پروتون است. وونا در سلولهای همه موجودات زنده ، از جمله در اولین تخمیر کننده های بی هوازی یافت می شود ، جایی که ATP در واکنشهای فسفوریلاسیون بستر سنتز می شود. سلولهای اولیه فرضی تمام انرژی را برای ضایعات فسفوریلاسیون بستر و ساختمانهای بیوسنتز بدخیم از بین بردند. کفایت ترکیبات ضروری ارگانیک از مرکز چشم انداز و چشم انداز محصولات پایان جنسی که در پشت مکانیسم تک پورت منفعل سرگردان هستند (شکل 26). در واقع سلولهای اولیه سلولهای کوچکی نیستند ، اما گلوله ها تنها با یک غشای اولیه از ظاهر بصری جدا می شوند. بدیهی است ، فرآیندهای حمل و نقل فعال ، که از انتقال ارتعاشی سخنرانی ها در برابر تمرکز شهرها جلوگیری می کند ، بول در اولین مراحل فرهنگ تکامل ضروری است.
به منظور ملاقات ، یک پمپ پروتون ته نشین ATP در سلول ها ایجاد شد و گلوله در CPM قرار گرفت. انرژی هیدرولیز ATP ، جذب ATPase ، برای پروتونهای vyshtovuvannya از سلولهای وسط پیروز بود. هیدرولیز یک مولکول ATP برای تولید تا انتقال 2 پروتون و ساقه به روشی از شیب پروتون الکتروشیمیایی غشایی. به صورت تجربی ، این مورد برای باکتری های اسید لاکتیک و کلستریدیوم نشان داده شده است ، که در آنها هیچ واکنشی وجود ندارد ، یا ATPase موضعی در CPM وجود ندارد ، به طوری که مولکول های ATP می توانند شکسته شوند ، که در هنگام تخمیر ایجاد می شود.
در چنین رتبه ای ، ATP vikorystannya برای ساقه H + روی غشا به صورت تکاملی حتی مکانیسم باستانی سلولین های پروکاریوتی است. مکانیسم Pіznіshe winyk سنتز ATP برای rakhunok H +. با توجه به تقاضای زیاد ، تغییر مستقیمی در مجموعه روباتیک پروتون ATPase ایجاد شد.
عملکرد معکوس ATPases پروتون در بی هوازی های اولیه شناخته شده است ، زیرا انرژی را در فرآیند تخمیر بازیابی می کنند. مشخص شد که بینایی در میان اسیدهای لاکتیک و اوسیک با باکتری های اسید لاکتیک و کلستریدیوم باید تا درجه درجه بندی پروتون در CPM تولید شود. در استرپتوکوک ها ، که تخمیر اسید لاکتیک سالم و هموفرمانتیو هستند ، اسید لاکتیک در سلول های آنیون تجمع می یابد ، که CPM عملاً غیرقابل نفوذ است. ترشح لاکتات از سلولها در فرآیند علائم الکترون خنثی با پروتونها جذب می شود (شکل 90). تصویری مشابه توسط ترویج می شود کلستریدیوم پاستوریانومهر چیزی که در سلولها و استات تجمع می یابد از طریق CPM به شکل جدا نشده عبور می کند. اسکیلکی همه طبقه متوسط غلظت اسیدهای لاکتیک و آلی روی لپه تخمیر در غذا ، اما نه در وسط جهان ، به دنبال کمک اضافی حاملان فعلی در غلظت ، قبر باشید. انتقال H + به منظور همدردی با لاکتات یا استات باید قبل از تولید در H + CPM انجام شود. با تجمع اسیدها در غلظت متوسط اسیدها ، گرادیان غلظت به تدریج کاهش می یابد ، در نتیجه درجه بندی پروتون ایجاد می شود ، پانسمان با اسیدهای قابل مشاهده از سلول ها تغییر می کند. در صورت غلظت بالای اسیدهای لاکتیک و اتزتوویک در وسط بازار ، H +، فقط با هیدرولیز ATP روی غشا رسوب می کند.
انرژی الکتروشیمیایی شیب پروتون ، که در فرآیند تخمیر در حضور اسید در طی فرآیند تخمیر رخ می دهد ، می تواند برای حمل و نقل در رودخانه های خاص و همچنین برای سنتز ATP مورد استفاده قرار گیرد ، که باعث می شود صدا ... دیدگاه انرژی برای راخونوک دیدن از مشتریان محصولات سرگردان می تواند قابل توجه باشد. در صورت تخمیر هموفرمانتیکال اسید لاکتیک ، پس از انجام pidrachunks ، می توان 30 درصد از انرژی مصرفی را اشغال کرد ، تا بتوان آن را بلعید. بنابراین ، در برخی از یوباکتریها ، که انرژی را در فرآیند تخمیر تشخیص می دهند ، ATP را می توان در واکنشهای فسفوریلاسیون بستر و علاوه بر این برای توسعه vict H +ویکتوری سنتز کرد ، که قرار است از ورود علائم محصول جلوگیری کند. در حال حاضر ، یوباکتریها با نوع تخمیر کننده اجباری نیروها نیز ممکن است دارای ATPases پروتون باشند که مستقیماً در هیدرولیز و سنتز ATP عمل می کنند ، یعنی گرگینه ها را به دو روش Catalyze می کنند:
ATP H +
نارشتی ، در برخی از باکتریهای بی هوازی اولیه سرگردان ، فعالیت ATP-synthase آشکار می شود ، که از قطعات کوتاه انتقال الکترونها در پشت یک نوار اضافی متصل به غشای حامل ها بدست می آید (div. Nizhche).
خرابی سیستم و انتقال دستگاه های الکترونیکی به O 2 در اولین بی هوازی ها
در واقع ، انتقال وسایل الکترونیکی اساس همه فرایندهای اکسیداتیو-حیاتی است. V انواع متفاوتشوموان ، نگاه کنید به فصل. 13 ، انتقال الکترونها (آب) از برخی مولکولهای آلی به مولکولهای آخر ، به توسعه NAD-dehhydrogenase بازگردید:
de DG - نوع دهیدروژناز ، برای انتقام از NAD مانند یک کوآنزیم ؛ A - یک مولکول گفتار آلی ، که به عنوان پذیرنده الکترون عمل می کند. مولکولهای NAD-H2 در متابولیسم سازنده فعال می شوند ، بنابراین از فرایندهای بیوسنتزی و همچنین در سیستم متابولیسم پرانرژی جلوگیری می کند و از سرنوشت "مشکل پذیرنده" ویروسی جلوگیری می کند. انتقال الکترونیکی به کل vypadku تا زمانی که انرژی otrimannya klitinoyu تولید نمی شود ، تنها در واکنشهای بستر فسفریله نقض نمی شود.
در برخی از توصیف های eubacteria ، انتقال مستقیم الکترون ها از آنزیم های اصلی NAD به O2 باید تا به روز رسانی بعدی داده شود:
OVER-H 2 + O 2 VER OVER + + H 2 O 2
اکسیداسیون دهیدروژنازهای NAD-fallow نیز در پشت فلاوپروتئین های میانی رخ می دهد ، که انتقال 1 ، 2 یا 4 الکترون به O2 و سپس تولید آنیون سوپراکسید ، پراکسید آب یا آب را کاتالیز می کند. O2 و H2O2 را می توان توسط آنزیم هایی که به طور کلی غذا را جمع آوری می کنند ، پخش کرد.
در بی هوازیهای هوازدنده ، مانند باکتری های اسید لاکتیک و deyaki clostridia ، فلاوپروتئین ، آنها نقش لانکای ثانویه اصلی را به عنوان بستر و اسید مولکولی ایفا می کنند. به عنوان مثال ، برای ایجاد ذهن بیهوشی در نتیجه تعقیب O2 از وسط ، برخی از سیستم ها می توانند جنایتکار باشند ، اما این بدان معنا نیست که این موضوع با انرژی مشتری دور ریخته می شود. تجدید O2 ، هنگامی که در نقش اکسیدازها ، یعنی آنزیم ها ، که بدون انتقال پیشینی الکترونها به رطوبت مولکولی ، فلاوپروتئین ها را معرفی می کنند ، من آن را "flavin dyhannya" می نامم. در اصل ، با فلاوین دیچانا ، انتقال دو الکترون به O2 وجود دارد. بنابراین ، در باکتری های اسید لاکتیک از جنس استرپتوکوک 90٪ O2 چسبیده به H2O2 تبدیل می شود.
نارشتی ، در برخی اوباکتریها ، اکسیدازهایی با طبیعت فلاوپروتئینی آشکار شده است که بسترهای اکسیده شده مستقیم ، به عنوان مثال ، اسیدهای پیروویک و لاکتیک را با ترش مولکولی کاتالیز می کند:
برنج غرق در مسیرهای حمل و نقل الکترونیکی برای مشارکت یک یا دو مورد در O2 - طولانی شدن واکنش ها در سیتوپلاسم سلول ، به عنوان مثال.
یک قلاب دوزی اساساً مهم در راه ریشه سیستم های انتقال الکترونیکی ، که برای تولید انرژی توسط سلول است ، به معرفی حامل های الکترونیکی به داخل غشا تبدیل شده است.
تشکیل دروازه ها با انتقال غشایی الکترون ها در سر بی هوازی
در کلاس اول بی هوازی ها ، شورت های کوتاهی برای انتقال الکترون ها وجود دارد ، به طوری که آنها می توانند پشت حامل های اضافی متصل به غشا قرار گیرند. در برخی موارد ، چنین انتقال بر روی حرکت پروتون ها در سراسر غشا قرار می گیرد و منجر به ایجاد H + و سنتز ATP می شود. یکی از موفق ترین همسران از این نوع ، سیستم فومارات ردوکتاز است که منجر به معرفی فومارات به سوکسینات می شود.
تجدید فومارات به سوکسینات را می توان به عنوان vicoristano برای اهداف آنابولیک (نیاز به سوکسینات برای سنتز تتراپیرول ها) ، حتی در فرایندهای کاتابولیک استفاده کرد. در آخرین راه حل ، همه اجزای واکنش می توانند متفاوت باشند ، و فقط برای پذیرش دستگاه های الکترونیکی (شکل 91 ، 4 ) ، یا اگر آن را در یک نوار متصل به غشا تغییر دهید (شکل 91 ، ب-د) برای ادای احترام واضح ، انتظار تولید سنتز ATP را نداشته باشید. پروتون Osvita بر روی غشاها هنگام انتقال الکترون ها به فومارات به منظور ذخیره در انبار و حامل های الکترونیکی roztashuvannya منتقل می شود.
اهداکنندگان الکترون برای تجدید فومارات می توانند به عنوان NAD-H2 ، لاکتات ، فرمت یا آب مولکولی عمل کنند ، از این طریق الکترون های پشت بسترهای اضافی دهیدروژنازهای خاص را می توان به حامل های متصل به غشا منتقل کرد (شکل 91 ، ب) حامل های متوسط در شناسایی پروتئین های FeS ، مناکینون و سیتوکروم ب، با این حال ، انتقال این نوع عدم لباس پوشیدن به رد انرژی روحانیت.
برای روشن کردن شیب پروتون در چند قطره ، یک اهدا کننده الکترونیکی و یک گیرنده الکترونیکی به انتهای غشا اضافه کنید. نیاز به الکترون در حامل ، محلی سازی در آخرین سمت غشا ، برای تولید تا پروتون های قابل مشاهده در وسط ، و تجدید فومارات در طرف دوم غشا روی سیتوپلاسم قرار می گیرد ، که طی آن پروتون ها قرار می گیرند. از طریق غشا منتقل می شود. تراز کردن شیب پروتون در پشت سنتز ATP کمکی منجر به سنتز ATP می شود (شکل 91 ، ب) همچنین ، دهنده و پذیرنده الکترونها در یک طرف غشا قرار دارند ، به طوری که شیب پروتون در همان روز هنگام انتقال الکترونها و روی فانوس شروع نمی شود ، فقط برای جایگزینی شمع حامل ها (شکل . 91 ، G) انتقال آب از طریق غشا از کینون ها کمک می گیرد. در ادامه جزئیات حمل و نقل الکترونیکی به سیتوکروم های موجود در غشا وصل می شود.
توانایی سنتز ATP در حین انتقال الکترون از NAD-H2 ، فرمت ، لاکتات ، H2 به فومارات بستگی به مقادیر پتانسیل های اکسیداتیو هدایت کننده و گیرنده سینتیک الکترون دارد.
عملکرد مشابه در سیستم کاتابولیسم بالینی ، تجدید فومارات به سوکسینات در تعدادی از یوباکتریها نشان داده شده است که در فرآیند تخمیر انرژی می گیرند. یکی از مراحل در راه ایجاد اسید پروپیونیک با تخمیر اسید پروپیونیک ، معرفی فومارات به سوکسینات است که کاتالیز کننده فومارات ردوکتاز است (شکل 54). فومارات ردوکتاز در کلستریدیا و استرپتوکوک اسید لاکتیک نیز شناخته شده است.
پانسمان ویدومی خوب با غشاء ، آنزیم سوکسینات دهیدروژناز کاتالیز شده در TCA اکسید سوکسینات به فومارات. هرگاه در واکنش فلاوین آندینوکلئوتید (FAD) پذیرفته شد ، بدون پیش فرض باید وارد لنز دیشال شود (شکل 92). نوسانات فومارات ردوکتاز و سوکسینات دهیدروژناز همان واکنش را ، هر چند در جهات مختلف ، از یک آنزیم مشابه کاتالیز می کنند. عفونت نشان داده شده است که واکنش قادر است با دانه های کوچک آنزیمی توسعه یابد. اطلاعات مربوط به آنها در ژن های مختلف یافت می شود. سنتز سوکسینات دهیدروژناز در ایروبیک و فومارات ردوکتاز - در ایروبیک فعال می شود.
کلستریدیوم استوژنیک سنتز استات را از CO2 و H2 نشان می دهد:
2CO 2 + 4H 2 CH 3 -COOH + 2H 2 O.
بوی تعفن می تواند در وسط شیمیایی رشد کند ، به طوری که می توانید به دلیل همان انرژی از H2 انتقام بگیرید. همچنین ، در بسیاری از ارگانیسم ها ، CO2 به استات ارتقا می یابد ، در سایر ضایعات با اکسید H2 یافت می شود ، اما با انرژی دارچین ارتباطی ندارد. مشخص شده است که در انتقال الکترون از H2 به CO 2 ، که منجر به سنتز استات می شود ، بخشی از فلاودوکسین ، مناهینون و سیتوکروم را انتخاب کنید. ب، T. E. حامل هایی از نوع مشابه در عملکرد سیستم فومارات-ردوکتاز.
در چنین رتبه ای ، در بین تعدادی از بی هوازی های اولیه ، که انرژی را در فرآیندهای سرگردان تشخیص می دهند ، یک اتصال کوتاه با یک غشاء ، یک لنز انتقال الکتریکی فرموله شد ، عملکرد چنین دستگاهی تا تایید شیب پروتون ، برای سنتز یک سنتز. از طریق رویت یک نوع پذیرنده انتهای خط الکترونها در ذهن بیهوشی ، توان خروجی در این نوع فرایندها کم است. با این حال ، اصول نوع جدید انرژی شکل گرفته است.
برای انتقال به انرژی vikorystannya نور ، لازم است مولکول های گیرنده نوری را بدست آورید و بخشی از آنها را به لنزهای باربری الکتریکی آشکار متصل کنید. چنین گیرنده های نوری - Mg -porphyrinis - بول ایجاد می شود. فتوسنتز از سیستم سیستم انتقال الکترونیکی چرخه ای ناشی از عکس شروع شد و به عنوان یک همراه در کیفیت انرژی dzherel ، تا مرحله اصلی ، مانند فرآیند سرگردان ، عمل می کرد. بازسازی موجودات اولیه فتوسنتز را می توان با وسیله نقلیه قطع کرد ، که می تواند برای بخشی از ATP سنتز شده در فرآیند انتقال دوار الکترون ها استفاده شود. در نتیجه ، ساختمان تجدید عکاسی مستقیم NAD + برای کل کسب و کار شکل گرفته است ، که نور حمل و نقل الکترونیکی غیرچرخه ای را تا نقطه عزیمت فراخوانده است. در دورتر ، یک دستگاه فتوسنتزی کامل با کیفیت اهدا کننده الکترون به vikorystannya vodi فراخوانده شد ، محصول جانبی آن تأیید ترش مولکولی بود.
در نتیجه دید فتوسنتزی O2 ، به نظر می رسد از نظر شیمیایی به عنوان یک عامل اکسید کننده فعال عمل می کند. در صورت ظهور O2 ، تعداد بیشتری پروکاریوت به توسعه مکانیسم ها منتقل می شود. برای برخی از افراد ، خط در انتها مستعد O2 است ، در نتیجه ، شکل بی هوازی با سطح رو به رشد هوادهی ایجاد می کند.
یک گام مهم در شکل گیری مکانیسم ترشح مولکولی در نقش گیرنده جنبشی الکترونها ، یک فرایند جانشین پرستاری است که برای رد قدرت مشتری ، نیروی پر انرژی زیادی را به خود اختصاص می دهد. در واقع ، انرژی زیادی وجود دارد که چگونه با جفت الکترون منتقل شده به صدا در بیاییم ، هم از نظر ماهیت دهنده ، هم از نظر ماهیت پذیرنده الکترون ها دراز بکشیم. به عنوان مثال ، پتانسیل اکسیداسیون NAD-H2 320 میلی ولت و اسید مولکولی +810 میلی ولت است. برای تأیید 1 مولکول ATP ، انتقال یک جفت الکترون با توجه به شیب الکتروشیمیایی ، با توجه به اختلاف بالقوه تقریباً 200 میلی ولت ضروری است.
برای vikorystannya O 2 در ظرفیت پذیرنده پایانی الکترونها در فرآیندهای مرتبط با کاهش انرژی متابولیک ، تبدیل الکترونیکی فتوسنتزی به دیکنی یکی از کمترین تاشدنی ها بود. در همان زمان ، لازم است که دهیدروژناز را به انتهای کم پتانسیل لانسوگا و سیتوکروم اکسیداز را به پایین ترین حالت ، که یک روش بدون میانگین O2 است ، اضافه کنید.
اساساً ، این گیاه تا ساقه سیستم آنزیمی برای تجدید چهار الکترونی O2 (سیتوکروم اکسیداز) ساخته شده است ، در این صورت هیچ محصول صنعتی سمی وجود نداشت.
فسفوریلات ، با توجه به انتقال الکترونها از بسترها در واکنشهای اکسیداتیو تیره ، فسفریلات اکسیداتیو نامیده می شود. توسعه مکانیسم فسفوریلاسیون اکسید امکان تولید بیشترین مقدار انرژی از بسترهای اکسیده شده را فراهم کرد.
در چنین رتبه ای ، ظهور ترش مولکولی باعث تکامل انواع جدیدی از زندگی در پرتو باکتری ها شده است ، که بر اساس رد انرژی برای فرایندهای فسفوریلاسیون اکسید است.
|
صفحه اصلی |
فاجعه Kisneva یک تغییر جهانی در انبار جو زمین است که در زمان دوران پروتروزوییک نزدیک به 2.4 میلیارد روبل شده است و نتیجه آن ظاهر یک ترش طبیعی در جو بود. در آن دوره ، ویژگی جو از اصلی به اکسیداتیو تغییر کرد. نظریه وینیکل در مورد فاجعه کیسنوو بر اساس ادای احترام
در جو اولیه زمین ، مولکولهای ترش رذیله ظاهر شد و او شخصیت خود را از اصلی تا اکسیداتیو به یاد آورد. برای 200 میلیون روبل ناقص ، غلظت ترش در فضای پروتروزوییک 15 برابر افزایش یافته است.
اعتراف در مورد فاجعه Kisnev بر اساس رشد رشد شراب به ماهیت تجمع کالاهای افتاده شکسته شد. از نظر بیولوژیکی ، ما باید در محل ترش شدید جو ، به اصطلاح نقطه پاستور ، زندگی کنیم ، به طوری که از کمی ترش در یک فضای مدرن به 01/0 نزدیک می شود. در سمت راست ، تنها با چنین رکود جوی موجودات زنده می توان از نتایج شریر تخمیر آنزیمی به اکسیداسیون موثرتر به مدت یک ساعت رسید. در دوران پروتروزوئیک ، نقطه پاستور نه تنها به آن نرسید ، بلکه کمی نوار بیولوژیکی طبیعی نیز به ارتعاش تکاملی راست اضافه شد - گسترش و توسعه گسترده تقریباً همه انواع سیارات زنده روی کره زمین.
پرورش دهندگان تماس های قابل توجهی در ترش ، در جو و هیدروسفر زمین نشان داده اند ، شیوه زندگی موجودات هوایی تک خط حفظ شده است ، که فقط می تواند در مستعمرات به اصطلاح ترش توسعه یابد. چرا ترشی در جو دوران پروتروزوییک اینقدر جوان شد؟ بر هیچ کس پوشیده نیست که رشد و باکتری های فتوسنتز کننده ، که در دوران آرکیه یافت شده اند ، سر بلندی یوگی هستند. اگر می خواهم دسته ای از ossyag توسط آنها چرخانده شود ، ترشی در جو و هیدروسفر کره زمین عملاً افزایش نمی یابد ، معلوم می شود که بلافاصله در اثر اکسیداسیون سنگ های داغ و گازهای جو آسیب می بیند. اگر تمام سنگهای سطحی و گازهای جو زمین اکسیده شده باشند ، گسل به گام موسلین در همان ویلیاد شروع به تجمع می کند. در مرحله پروتروزوییک تاریخ زمین ، غلظت اسیدیته در نتیجه زندگی باکتری ها در تاریخ زمین 1 of از نوبت عوضی... به جای دی اکسید کربن ، کاهش تدریجی ویتراته دی اکسید کربن در فرآیند فتوسنتز جلبک ها.
Otzhe ، فاجعه Kisneva میراث کوچک و باشکوهی برای تکامل موجودات زنده است. اتمسفر و هیدروسفر سیاره ما به جای کلماتی که در زمین کمتر از فضا هستند ، از کلمات سبک و پرنده تشکیل شده است. هنگامی که زمین شکل گرفت ، کیست ها به انبار رودخانه های جامد منتقل شدند ، پاشیدن نیتروژن - به نیترید و کیسن - به اکسیدهای فلزی.
در فرآیند فعالیت آتشفشانی فعال در دوره پیش زمین شناسی تاریخ زمین ، بازالتهای ذوب شده ، بخار و گازهای گوشته فوقانی در حال رشد بودند. Yak به آخرین نشان داده شد ، آتشفشان های شادآنها بخار آب و همچنین گاز دی اکسید کربن ، کلر ، متان و سایر اجزا را مشاهده می کنند. اگر در دمای بالاتر ، بخار به اتمسفر منتقل می شود ، که به آن اسید dimis - spoluca sirki ، اسید بوریک و نمک های آمونیوم می گویند. از نظر ظاهری ، جو اولیه زمین در نتیجه گاز زدایی از گوشته شکل گرفت و اساس آن گاز دی اکسید کربن ، آب ، آمونیاک و متان بود.
برای ارزیابی تغییرات ایجاد شده در جو و هیدروسفره سیاره سیاه در دوران پروتروزوییک ، لازم است به انبار جو اولیه مراجعه کنیم. ویوچنیا به جای پیازهای گاز در کوارتزیت باستانی که در سازند کورومکان سپر آلدان یافت شد به ما اجازه داد تا انبار جو اولیه زمین را روشن کنیم.
در پیازهای سیچ ، هیچ نهالی وجود ندارد ، در انبارهای آنها 60 درصد وام دی اکسید کربن و تقریبا 35 درصد آب ، اکسید آب ، آمونیاک و اسید کم نور است. بدیهی است که برخی اجزا در هنگام گاز زدایی گدازه ها به سطح زمین آمده و در چنین مرتبه ای ، پوسته ای بسیار نازک ایجاد کرده اند. دمای چنین فضایی در نزدیکی سطح سیاره در وسط 15 درجه سانتیگراد شد. بخار آب گازهای آتشفشانی متراکم شده و به آب تبدیل شد. هیدروسفر زمین به این شکل شکل گرفت. در کره زمین ، به محض ایجاد اقیانوس اولیه ، کدی عبور کرد ، به آب رسید ، انبارهایی از قسمتهای گازهای آتشفشانی. در مرحله پیش زمین شناسی و باستان شناسی تاریخ سیاره ، کمبود آب در اقیانوس ها ؛ تیلکی در رقیب پروتروزوئیک به قله های نارستی اقیانوس دوسیاگ آخ.  تعداد زیادی از گازهای آتشفشانی جوی ، که در نزدیکی آب واقع شده اند ، انبار اقیانوس اولیه برای زمان نژادهای دریایی هستند که بر روی سطح زمین و در پایین دریا ، ویرانه های بیداری خواب آلود
تعداد زیادی از گازهای آتشفشانی جوی ، که در نزدیکی آب واقع شده اند ، انبار اقیانوس اولیه برای زمان نژادهای دریایی هستند که بر روی سطح زمین و در پایین دریا ، ویرانه های بیداری خواب آلود
یاک در جو و هیدروسفر زمین ظاهر می شود؟ مهم این است که مولکولها با استفاده از بخش کوچکی از مولکولهای بخار آب به همراه جزء خشن نوشیدنی خواب آور ایجاد شوند. اعتراض در فرایند واکنش نشان داده می شود ، گناه حتی ناچیز خواهد بود ، زیرا گاز خود را با ویپرومینوانه های ماوراء بنفش پوشانده و مولکول های آب را تقسیم می کند.
در چنین رتبه ای ، تغییر عنصر شیمیایی لازم برای زندگی در فضایی از archaea bulo حداقل است - nabagato کمتر از هزار تن rivnya روزانه است. در عین حال ، عملاً تمام مولکولهای تشکیل شده به سرعت بر روی گازهای اکسیده شده اتمسفر شیشه ای شدند. جو ظریف اولیه در ساعتی از روز نمی تواند این سیاره را از ابتدای توسعه بیولوژیکی زمین از شر vipprominuvannya Sontsya سخت بگیرد.
تا گوش Proterozoic ، کمی آب در زمین prodovzhuvalo zbilshuvatisya - که خود را به عنوان یک اقیانوس مقدس تثبیت کرده است. اما در همان زمان ، غلظت دی اکسید کربن در جو اولیه پروتروزوییک کاهش یافت. و تغییر ترش در جو و هیدروسفر کره زمین بسیار کم شد - فقط 1 of از فصل جاری.
انتقال می یابد ، در کل دوره در گوشته زمین ، 4-6 deposit از رسوب فلز گرفته می شود ، که نقش خاک رس سخت فشرده است. یک عنصر شیمیایی سه ظرفیتی کامل ، بدون تسلط در آب ، قبل از محاصره و تجمع همزمان با سیلیس از بارهای باشکوه سنگ معدن طلا که امسال شاهد آن هستیم. بنابراین ، در جو اولیه پروتوزوئیک سیاره ما در انبار اصلی ، نیتروژن با افزودن کمی بخار آب ، آرگون ، دی اکسید کربن و اسید محروم شد. فاجعه Kisne به زندگی بزرگی در Proterozoic تبدیل شد. از نام زمین در تاریخ زمین ، انقلاب گذشت ، که در آن 2.4 میلیارد روبل شده است. جو سیاره ما در ساعت tsei شبیه ترشی در مقیاس بزرگ خواهد بود.
فاجعه Kisneva (انقلاب Kisneva) - تغییر جهانی در انبار جوکه شبیه یک آرکئا شد - یک لپه پروتوزوئیک، نزدیک به 2.4 میلیارد روبل تا آن زمان (دوره سیدریت) نتیجه فاجعه Kisnevoy ظاهر شدن در انبار جو a بود ترشі تغییر شخصیت zagalny جو از اصلی به اکسیداتیو. اعتراف در مورد فاجعه Kisnev بر اساس رشد رشد شراب به ماهیت تجمع کالاهای افتاده شکسته شد.
تا تغییرات قابل توجهی در جو موجود در جو ، حتی در جدیدترین شکل زندگی بول بی هوازی، Tobto مبادله کلمات در اشکال زنده ، پس از سقوط به اشکال آخوندی dikhannya ، به عنوان آنها ترش vimagali نیست. دسترسی به ترش در تعداد زیادی از دندانها برای تعداد زیادی از باکتریهای بی هوازی است ، زیرا در این زمان بخش بزرگی از موجودات زنده بر روی زمین وجود دارد. راشتا عمر یک بول را تشکیل می دهد ، یا قابل اکسیداسیون و پوسیدگی دندان نیست ، یا چرخه زندگی خود را در وسط بدون اسیدیته انجام می دهند.
تجمع O2 در جو زمین:
1. (3.85-2.45 میلیارد روبل) - O 2 انجام نشده است
2. (2.45-1.85 میلیارد روبل به آن) O 2 انجام شد ، آل پوشیده از اقیانوس و سنگهای بستر دریا
3. (1.85-0.85 میلیارد rok_v that) O 2 از اقیانوس خارج می شود ، آلی با سنگهای سنگی اکسید شده در خشکی و هنگام ایجاد توپ ازن
4. (0.85-0.54 میلیارد روبل)
5. (0.54 میلیارد روبل پیش-تا به امروز) مخازن O 2 ذخیره و تعمیر می شوند تا در جو تجمع پیدا کنند
ذخیره اولیه جو توسط پروتروزوییک
انبار دقیق جو اولیه زمین در روز ناراحتی امسال ، اعتراض به دانش ، زیرا در نتیجه گاز زدایی از گوشته شکل گرفته بود و کاملاً متفاوت بود. اساس її شد گاز دی اکسید کربن, سیروددن, آمیاک, متان... در مورد دارچین سگو کاهش دهید:
- رسوب بدون اکسیداسیون ، که به وضوح در سطح ایجاد شده است (به عنوان مثال ، یک سنگریزه کوچک از ناپایدار تا ترش پیریتو);
- مشاهده گونه های قابل توجه ترشل ژرل و سایر اکسید کننده ها.
- vivchennya dzherels بالقوه جو اولیه (گازهای آتشفشانی ، انبار اولین اجرام آسمانی).
علل فاجعه ترش
Є یک ژرل مهم ترش مولکولی є بیوسفر ، به طور دقیق تر ، فتوسنتز کننده organizmi ظاهر شدن روی لپه زیست کره ، فتوسنتز می کند آرکی باکتری هاآنها بوسه ای را چرخاندند ، که عملاً بلافاصله به یک ژیرسکیخ پورود اکسیده ، rozchinenih spoluk و گازهای جو تبدیل شد. غلظت ویسوکا به صورت محلی ، در بین ، گسترش یافت مواد باکتریایی(T. صدا "Kisnevi kisheni"). بدین منظور ، هم سنگهای سطحی و هم گازهای اتمسفر تبدیل به اکسیداسیون شده اند و هم رطوبت در بیابان در اتمسفر جمع شده است.
علاوه بر این ، یکی از عوامل جوانی که در تغییر روح میکروبی تحت تأثیر قرار گرفته است ، تغییر شیمی به اقیانوس است. بنابراین ، برای یک z فرضیه هاعملکرد حصیر باکتریایی باستانی را می توان با کاهش غلظت خفه کرد نیکلمن نقش مهمی در متانوژنز... کاهش غلظت این سخنرانی ها و سخنرانی های دیگر می تواند به دلیل از دست دادن فعالیت های آتشفشانی باشد.
آثار فاجعه کیسنوو
زیست کره
Oskіlki قسمت اعظم بدن را در آن ساعت گاو نر بی هوازی، nsnuvati بدون فکر با غلظت قابل توجهی از ترش ، تغییر جهانی در spilot دیده می شود: بی هوازیروح تغییر کرده است ایروبیک، بین لیش قبلی "kishenami ترش" ؛ بی هوازیخوب spіlnoti ، navpaki ، vyavilsya vitisnenі در " بی هوازی kishenі "(از نظر مجازی به نظر می رسد ،" جو به ناویوریت تبدیل شد "). ظاهر ترش مولکولی در جو منجر به شکل گیری آن شد صفحه ازن، بطور قابل توجهی محوطه زیست کره را گسترش داده و به وسیل وسیعتر انرژی وسیع تری را فرا می خواند بی هوازی) Kisnevoy dikhannya.
لیتوسفر
در نتیجه فاجعه اسیدی ، عملاً همه دگرگونі نژادهای محاصره، برای تبدیل شدن به بخش بزرگی از سرخک زمین ، є با اکسیداسیون.
Сідер (в گردوσίδηρος - zalizo) - دوره زمین شناسی ، بخشی پالئوپروتروزوییک... بازه ساعتی از 2.5 تا 2.3 میلیارد سال پیش. Datuvannya صرفاً زمانی است ، نه بر اساس لایهنگاری.
در گوش دوره خاصی از حمله حاوی آهننسل x اگر بوی تعفن در ذهن شکل می گرفت ، اگر رشد بی هوازی آبتغییرات را ریشه یابی کرد mussen، Yakiy ، zmіshuyuchis z zlіzom ، تأیید مگنتیت(Fe 3 O 4 ، اکسید زالیزا) فرایند Tsey پاکسازی zalizo اقیانوس ها... در کیسه kintsevoy ، اگر اقیانوس ها آن را فشار داده اند ، روند پیوند تا انتها جو، Yaku mi maєmo در این روز.
جو
در نتیجه تغییر شیمی جو فاجعه اسید ، تغییر در فعالیت شیمی ، توپ ازن تشکیل شد ، اثر گلخانه ای ... میراث یاک ، سیاره وارد عصر شد هورون یخی.
guronske zledeninnya
مطالب از ویکی پدیا
یخی Guronske - یافت شده و بی اهمیت ترین یخ زمین. سلطنت کرد و در عصر پایان یافت پالئوپروتروزوییک، Trivalo نزدیک به 300 میلیون راکی است.
باعث یخ زدگی شود
1. علت اصلی گاو یخی Huronian فاجعه کیسناوا، هنگامی که جو زمین ، تعداد زیادی وجود داشت ترش، متزلزل فتوسنتزموجودات زنده متان، Yaky قبلاً ، به دلیل حضور زیاد در جو و افزودن موارد اصلی در اثر گلخانه ای، پس از ملاقات با ترش و ایجاد مجدد در گاز دی اکسید کربنو آب. Zmini به انبار جو ، به جذابیت خاص خود ، به سرعت افزایش یافت متانوژن، Scho viklikalo dodatkove سطح متان را کاهش می دهد.
2. مقیاس عظیم و بی اهمیتی یخ Huron را می توان به خوبی به اصطلاح گره زد. تناقض سنتسیا جوان ضعیف.
3. نظریه "زمین-اسنیژوک" (انگلیسی زمین گلوله برفی) - فرضیه ، کمک هزینه Шо، Шо زمینگلوله پوشانده شده است یخدر بخش kriogeniyі اداکارعادت زنانه نئوپروتروزوییک eri، і، احتمالاً، در دوران زمین شناسی іnshі. خنک کننده در دی اکسید کربن C O 2 در اقیانوس ها و تبدیل آن به بخار Ca C O 3 را توضیح می دهد.
4. رشد قاره ها به شکل فوق تماس رودینیا و توسعه سپر kryzhany ، مشابه قطب جنوب.